
記住我
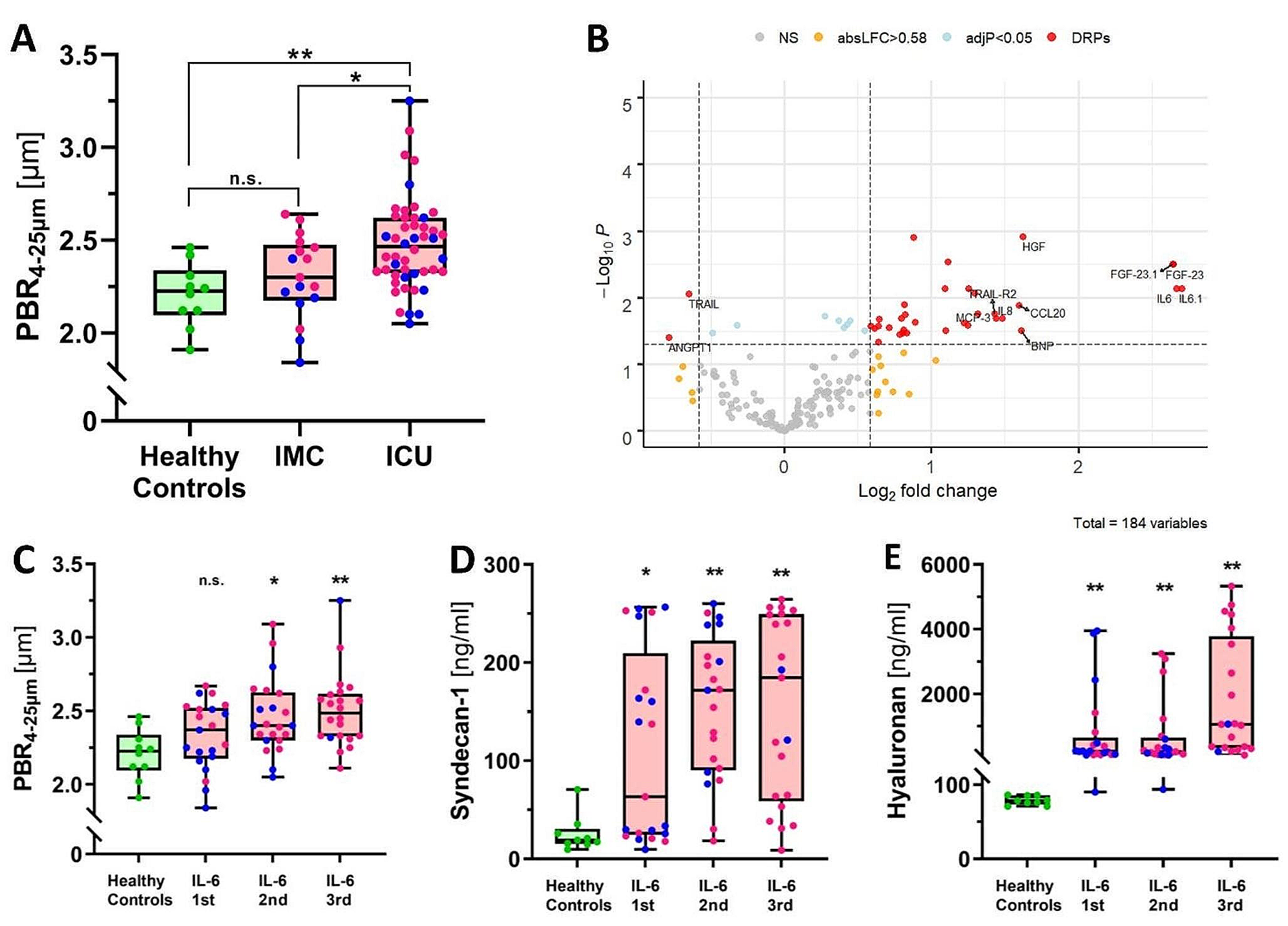


To model EndMT, we exposed ECFCs to oxLDL to study endothelial progenitors’ dysfunction and trans-differentiation [13] (Fig. 1A). Expectedly, an elongated and fusiform cell shape could be observed in sub-clusters of oxLDL treated ECFCs suggestive of EndMT (Fig. 1B). The changes in the expression of cell surface markers associated with oxLDL exposure were assessed by flow cytometry analysis. After 5 days of continuous oxLDL treatment, a significant and concentration-dependent decrease in the expression of CD34, a key endothelial marker, was noticed in ECFCs treated with oxLDL. Flow cytometry analysis showed 2.4 and 3.2-fold reduction in the percentage of CD34 + cells in ECFCs treated with 25 and 50 µg/mL of oxLDL respectively (p = 0.0001 vs. Veh, Fig. 1C and D). Additionally, CD90 (THY1), a key mesenchymal marker, was used to examine the gain of mesenchymal phenotype [14]. Conversely, 1.5 to 2.2% of ECFCs began to express CD90 at 25 and 50 µg/mL of oxLDL treatment respectively (p < 0.0001 vs. Veh, Fig. 1E and F). We confirmed this finding by immunostaining (Fig. 1G and H). Overall, these features validated the use of oxLDL as an established model of EndMT.
Fig. 1
Exposure to oxLDL results in progenitor dysfunction and EndMT-associated changes in ECFCs. (A) Schematic representation of experimental timeline and design. ECFCs were treated with vehicle or oxLDL at 3 different concentrations (12.5, 25 and 50 µg/mL) for 5-days. Cellular changes were assessed by FACS and single-cell colony forming assay. (B) Morphological differences can be observed in culture after a 5-day oxLDL treatment period. (C-F) Flow cytometry analysis of endothelial marker CD34 and mesenchymal marker CD90 on oxLDL treated ECFCs. (vs. vehicle; n = 5 ECFCs isolated from biologically independent donors; mean ± SD; p value was calculated by two-way ANOVA with multiple comparison to Vehicle) (G and H) Immunofluorescent staining of CD90 on oxLDL treated ECFCs (vs. vehicle; n = 5 ECFCs isolated from biologically independent donors; mean ± SD; p value was calculated by one-way ANOVA with multiple comparison to Vehicle). (I) ECFCs were then FACS sorted at a single-cell level for the examination of proliferative potential (vs. vehicle; n = 5 ECFCs isolated from biologically independent donors; mean ± SD; p value was calculated by two-way ANOVA with multiple comparison to Vehicle) (J and K) Morphological assessment of CD90 + and CD90- oxLDL treated ECFCs single-cell sorted and cultured for 7 days without oxLDL
A single-cell colony formation assay was performed to determine the functional consequence of oxLDL treatment on ECFC self-renewal. ECFCs were treated with 25 µg/mL of oxLDL for 5 consecutive days and then sorted by FACS, plated 1 cell per well in endothelial cell growth medium-2 (EGM2) without oxLDL. Each flow-sorted cell was allowed to proliferate over 14 days. A systematic and prospective classification process was devised to categorize types of colonies formed by individual cells. A single-sorted ECFC is able to produce one of three types of colonies: a high proliferative potential (HPP) colony (> 500 cells at 14 days), a low proliferative potential (LPP) colony (> 250–500 cells), or an endothelial cluster (< 50 cells) [4]. Sorted ECFCs within the vehicle treatment group gave rise to a significant proportion (69 to 78% of the total number of wells containing cells) of HPP colonies versus the oxLDL treatment group, which comparatively demonstrated an 8-fold decrease (p < 0.0001 vs. Veh HPP, n = 5). Conversely, 71 to 86% of the colonies formed within the oxLDL group were determined as endothelial clusters, indicating a loss of self-renewal and proliferative capabilities during the 5-day exposure to oxLDL (p < 0.0001 vs. Veh EC, n = 5, Fig. 1I).
We noted a discordance in the percentage of ECFCs with reduced CD34 expression versus the small, although significant percentage of CD90 positives ECFCs suggestive of a transitional phenotype of ECFCs at an intermediate stage of trans-differentiation in the EndMT process. To evaluate the reversibility of the phenotype, we sorted oxLDL (25 µg/mL) treated ECFCs using CD90 marker and assessed their colony morphological changes in EGM2 without oxLDL at the single cell level (Fig. 1J). After 7 days, CD90 + ECFCs formed a mesenchymal spindle-shaped morphology. In contrast, CD90- colonies maintained a classical endothelial cobblestone-like morphology (Fig. 1K). However, following a 14-day culture in EGM2 medium, all cells regained an endothelial phenotype and exhibited a loss of CD90 surface marker expression (Supplemental Fig. 1A-C). This suggested a transition phase of reduced endothelial markers where endothelial morphology could be recovered and a more advanced mesenchymal phenotype with CD90 expression where the transition to mesenchymal phenotype was more pronounced that could also recover after longer time.
EndMT dramatically alters chromatin accessibility and gene expression in ECFCsThe established and irreversible mesenchymal phenotype and loss of self-renewal pointed towards epigenetic alterations, prompting us to evaluate gene expression and chromatin accessibility changes. We assessed transcriptomic changes of oxLDL treated ECFCs (25 µg/mL, 5-days) via bulk RNA sequencing (n = 4 ECFC from different donors per group). The oxLDL treatment resulted in the downregulation of 3128 genes and upregulation of 2554 genes (p < 0.05 after correction for multiple testing, Fig. 2A). Key attributes of the endothelium such as CDH5, DLL4 and CD34 expression were downregulated and the expression of classical mesenchymal genes such as THY1(CD90), SNAI, RUNX2, BMP4 and TGFβ2 as well as a large number of extracellular matrix genes were upregulated in ECFCs exposed to oxLDL. Several known molecular pathways involved in EndMT, including the epithelial to mesenchymal transition and TGF-beta signalling pathway were identified to be significantly altered by exposure to oxLDL [15,16,17,18]. This was further confirmed by GSEA (Gene Set Enrichment Analysis) showing over-representation of Epithelial to Mesenchymal Transition and TGFbeta signalling genes among those upregulated in ECFCs exposed to oxLDL (Fig. 2B and C). This analysis also revealed significant changes in expression of genes associated with other pathways such as hypoxia or TNFalpha signalling (Supplemental Fig. 2A and B). We identified SOX9 among the top genes increased after oxLDL treatment (Fig. 2A), a chondrogenic/stem cell factor which has been previously reported to be expressed in murine endothelial progenitors, with its loss associated with endothelial differentiation as well as attenuated EndMT within the wound healing context in mice [10].
Fig. 2
Bulk RNA and ATAC-sequencing analysis between vehicle and oxLDL-treated ECFC groups display a significant difference in gene expression and chromatin landscape. (A) Volcano plot of differentially expressed genes between vehicle and oxLDL (5-days, 25 µg/mL) treated ECFCs. N = 4 biological replicates per condition. Blue dots signify significantly down-regulated genes and red dots represents significantly up-regulated genes in oxLDL treated ECFCs compared to vehicle. (B) Gene Set Enrichment Analysis (GSEA) plot for oxLDL-enriched hallmark Epithelial to Mesenchymal Transition and TGFbeta signaling (Molecular Signatures Database, MSigDB) showing the profile of the running Enrichment Score (ES) and positions of gene set members on the rank ordered list. NES (top), and FDR q value (bottom rows) are indicated on GSEA plot. (C) Heatmap of the top 15 differentially expressed genes from the EMT and TGFbeta pathway gene sets. Expression values are represented as colours and range from red (high expression) to dark blue (lowest expression). (D) Heatmap profile of global enrichment for ATAC-seq peaks demonstrating chromatin accessibility at genome-wide transcription start sites (TSS) ± 1 kb and at up-regulated gene promoters ± 1 kb in vehicle and oxLDL treated ECFCs. The blue color intensity reflects the level of peak enrichment. Each condition has 4 biological replicates. (E) Pathway analysis for the differential ATAC-peaks (N up-regulated after oxLDL treatment at FDR < 0.05 = 3633). (F) HOMER analysis of SOX9 motifs enrichment. (G) Permutation-based strategy using ENCODE dNase-seq footprinting data. Motifs for selected TFs (SOX9, GLI1, ZEB1 and CTCF) were intersected with treatment-dependent open chromatin peaks. The observed number of intersected motifs (red diamonds) was compared to null background sets (boxplots). (H) Pie charts graph representing part of the whole number of open chromatin peaks overlapping promoter region (within 5 kb) of up-regulated genes in oxLDL treated ECFCs and containing one or more SOX9 motifs (red)
To assess the extent of oxLDL induced EndMT in ECFCs, we used a previously published bulk RNA sequencing dataset comparing human primary meso-endothelial bipotential placental cells that gave rise to both ECFCs and mesenchymal stem cells (MSC) in culture [19]. Additional transcriptional analysis was performed by cross comparison between oxLDL vs. Veh and ECFC precursors vs. MSC precursors datasets. A large proportion of differentially expressed genes (DEG) were shared between these two datasets (Supplemental Fig. 2C and D). Upon further pathway analysis of shared genes upregulated in ECFC precursors and downregulated in oxLDL treated ECFCs, we observed shared endothelial-specific pathways including “cell-cell adherent junction” and “PECAM1 interaction” strongly supporting the loss of endothelial phenotype of ECFC upon oxLDL exposure. Conversely, “collagen deposition”, “ECM organisation” and “TGF-beta signalling” pathways were shared between genes upregulated in MSC precursors and oxLDL treated ECFCs supporting the transition of ECFCs exposed to oxLDL towards a mesenchymal phenotype (Supplemental Fig. 2C and D). In addition, GSEA confirmed enrichment of fibroblast signatures and loss of fetal vascular endothelial cells signatures in the oxLDL treated cells (Supplemental Fig. 2E-H). Overall, gene expression changes occurring in ECFCs upon oxLDL exposure largely recapitulated features of EndMT.
We next asked if transcriptional changes in ECFCs associated with oxLDL driven EndMT were associated with modifications in chromatin accessibility and performed ATACseq analysis (n = 3 for oxLDL treatment and n = 4 for vehicle). The treatment by oxLDL led to significantly altered chromatin accessibility in ECFCs that clustered based on oxLDL treatment (Supplemental Fig. 3A and B). Peaks were distributed mostly around annotated gene promoters and introns (Supplemental Fig. 3C). As expected, ATAC-seq signal was enriched at transcriptional start sites (Supplemental Fig. 3D).
We identified 3633 significantly enriched peaks in the oxLDL compared to the vehicle samples (defined as FDR ≤ 0.05, and fold change > 1), and 30 regions that were more accessible in the vehicle compared to the treatment (fold change < -1). The analysis of the enrichment for ATAC-seq peaks at genome-wide regions surrounding transcription start sites (± 1 kb; TSS) showed that oxLDL treatment induced a global increase in chromatin accessibility (Fig. 2D). This list was narrowed down by focusing on genes with increased expression upon oxLDL exposure from our RNA-sequencing results, revealing a correlation in chromatin accessibility at these genes’ promoters. Looking at enriched pathways after treatment among the differential ATAC-peaks (N up-regulated at FDR < 0.05 = 3633) and assuming that the nearest gene to the peak is a target gene, we could identify “extracellular matrix organisation” and “collagen formation” that were reminiscent of the “EMT pathways” enriched in RNAseq results (Fig. 2E). Interestingly, key signalling pathways associated with endothelial cells were also identified such as VEGF signalling. This highlights that ECFCs undergoing EndMT maintain an open chromatin allowing the potential re-expression of endothelial genes as observed in the CD90- fraction of ECFCs.
We next looked at transcription factor binding motifs in oxLDL treatment-dependent open chromatin. HOMER analysis showed that peaks upregulated upon oxLDL exposure were enriched for binding motifs of AP1-cJUN and CTCF (Supplemental Fig. 3G). We then asked if the changes observed in open chromatin could reflect classical transcription factors involved in EndMT. We compared SOX9, CTCF, GLI1 or ZEB1 motifs in the oxLDL dependent open chromatin using permutation analysis to reveal a greater overrepresentation of SOX9 motifs as compared to other transcription factors (Fig. 2G). Indeed, motifs associated with SOX9 activity (Fig. 2F) demonstrated a 1.25-fold increase in oxLDL-dependent open chromatin regions. Among the treatment-dependent peaks (N = 3633), 847 (23%) contained one or more SOX9 motif. This was 1.7-fold greater than what would be expected by chance, based on a random sampling strategy.
When examining genes with upregulated expression identified by RNAseq in oxLDL treatment, 95 were found to have an oxLDL-dependent open chromatin peak in proximity to their promoter region (within 5 kb), and 16 of those genes contained a SOX9 binding motif (Fig. 2H). This possibly reflects key genes where SOX9 is more directly responsible for the overexpression. Twelve of them (RASL11B, CRYAB, IL7R, DCN, FMN1, HES1, IRF6, TNFSF10, PDE4B, LRCH2, AHCYL1 and MNT) are associated with Atherosclerosis (a condition where EndMT is known to play a role) according to the CTD Gene-Disease Associations dataset and the gene set enrichment analysis (Supplemental Fig. 3E). Finally, vascular smooth muscle and myofibroblasts cell type signatures were found to be associated with these 16 genes containing a SOX9 binding motif (Supplemental Fig. 3F).
Together, these observations demonstrate that in vitro exposure of ECFCs to oxLDL increases chromatin accessibility leading to the expression of a transcriptional program allowing mesenchymal transition associated with reduced expression of endothelial genes. Among transcription factors potentially driving these chromatin accessibility changes, SOX9 could play an important role given the over-representation of its binding domains.
To further understand and validate the molecular kinetics of oxLDL treatment and associated EndMT changes in ECFCs, the key genes identified from the RNA-seq were examined at various timepoints at mRNA and protein levels (Fig. 3A). Firstly, SOX9 mRNA expression levels were assessed at various timepoints and oxLDL concentrations compared to respective time-matched vehicle control. Exposure to 25 µg/mL oxLDL resulted in the significant increase of SOX9 expression only by day 5 (p < 0.0001 vs. Veh 5-days, Fig. 3N). Immunoblotting and immunofluorescent staining of SOX9 further supported this increase at the protein level (p = 0.0030 vs. Veh, p = 0.0286 vs. Veh respectively, Fig. 3P-S). oxLDL treatment (25 µg/mL) resulted in a 3 to 6-fold increase in the expression of mesenchymal markers SNAI1 and FSP-1, as well as osteogenic marker RUNX2 (p < 0.0174 vs. Veh, Fig. 3B-D), with increasing expression occurring mostly from D5 of exposure to oxLDL treatment mirroring the expression of SOX9. Consistently, when using immunostaining, FSP-1 and RUNX2 were also expressed at higher levels in oxLDL treated ECFCs compared to vehicle group (p = 0.0022 vs. Veh, Fig. 3H-K). The observed increase of RUNX2 expression at protein level was confirmed by Western Blot (p = 0.0079 vs. Veh, Fig. 3L and M). Furthermore, the expression of the endothelial adhesion gene, CDH5 and those involved in the Notch signalling pathway (DLL4, HEY1) were significantly reduced in the oxLDL treated group (p < 0.007 vs. Veh, Fig. 3E-G). However, in the case of these genes the changes in expression occurred more continuously over time and before D5.
Fig. 3
oxLDL-treated ECFCs lose the expression of endothelial cell-specific genes and initiate the expression of mesenchymal cell-specific genes. (A) Schematic representation of treatment timeline and oxLDL dose. (B-G) mRNA expression of mesenchymal and endothelial genes after 25 µg/mL oxLDL treatment at varied timepoints. (vs. respective timepoint vehicle; n = 5; 5 ECFCs isolated from biologically independent donors; mean ± SD; p value was calculated by one-way ANOVA with multiple comparison to Vehicle). (H-K) Immunofluorescent quantification of FSP-1 and RUNX2 protein expression in ECFCs exposed to 25 µg/mL oxLDL for 5 days (vs. vehicle; n = 5; 5 ECFCs isolated from biologically independent donors; mean ± SD; p value was calculated by Mann–Whitney U test to Vehicle). (L and M) Western blot analysis of RUNX2 expression in ECFCs treated with 25 µg/mL (N and O) SOX9 mRNA expression in ECFCs at specific timepoints of oxLDL exposure and oxLDL concentrations. (P-S) Immunofluorescent quantification and western blot analysis of SOX9 expression in ECFCs treated with 25 µg/mL oxLDL for 5 days (vs. vehicle; n = 5; 5 ECFCs isolated from biologically independent donors; mean ± SD; p value was calculated by Mann–Whitney U test to Vehicle)
SOX9 shRNA knockdown attenuates oxLDL induced progenitor dysfunction and EndMT whereas SOX9 overexpression mimics EndMTTo further investigate the role of SOX9 in oxLDL-induced progenitor dysfunction and EndMT, transduced ECFCs expressing an shRNA against SOX9 upon tetracycline induction were generated from 2 different donors, here termed ECFC [9]. Integration and expression of shSOX9 was first validated using immunoblotting of the TetR repressor (Supplementary Fig. 4A). Also, the impact on SOX9 expression was measured using a various dose of tetracyclin demonstrating about 50% reduction at protein and RNA level (Supplementary Fig. 4B and C). Our data showed that EndMT-associated genes identified within the RNA sequencing to be regulated by oxLDL treatment SNAI1, FSP-1, CDH5 and DLL4 were not affected by the knockdown of SOX9 at baseline in the absence of oxLDL (Fig. 4A-F). Interestingly, within the vehicle treated group, the expression of SOX9 was not affected by the knockdown, consistent with its low/negligible expression at the basal level. SOX9 expression was induced (15-fold increased) upon oxLDL exposure (5 days, 25 µg/mL, p < 0.0001). Upon silencing, the increase in SOX9 expression after oxLDL exposure was reduced (4-fold only) and not significantly different from vehicle, suggesting effective blockade of SOX9 expression by the shRNA. Strikingly, after oxLDL exposure, SOX9 silencing was associated with a near-complete restoration of expression of endothelial genes (CDH5, DLL4) significantly reduced in the ECFCshScr. Conversely, the silencing of SOX9 strongly inhibited the aberrant expression of other mesenchymal genes induced by oxLDL treatment such as SNAI1 and RUNX2 and a partial but significant (66% reduction) attenuation for FSP1 within ECFC [9] compared to control ECFCshScr (Fig. 4A-F). In parallel, the flow cytometry analysis for CD34 and CD90 proteins was performed to determine whether this change in gene expression could influence the expression of key CD markers. While the CD34 + population was reduced upon oxLDL treatment, this was only partly restored by SOX9 silencing (p = 0.0425, Fig. 4 G and H). In contrast, the increased number of CD90 + cells after oxLDL treatment was effectively reversed in the ECFC [9] group (Fig. 4 I and J). These lines of evidence appear to be indicative of an attenuation of oxLDL-induced EndMT upon SOX9 silencing.
Fig. 4
SOX9 shRNA silencing attenuates oxLDL-induced progenitor dysfunction and EndMT. (A-F) Mesenchymal and endothelial mRNA expression assessed by qPCR. (G-J) FACS analysis of endothelial marker CD34 and mesenchymal marker CD90 expression of ECFCshScr and ECFCshSox9 exposed to 25 µg/mL oxLDL. (K and L) Capillary tube formation assessment of ECFCshScr and ECFCshSox9 exposed to 25 µg/mL oxLDL. (M and N) Analysis of ECFC motility by scratch assay. Quantification of the scratched area invaded by ECFCshScr and ECFCshSox9 treated with vehicle or oxLDL for 5 day within a 24-hour time-lapse (O and P) 3D collagen matrix gel contraction assay of ECFCshScr and ECFCshSox9 treated with vehicle or oxLDL, images taken at 24 h and 72 h post seeding. (Q) ECFC proliferative capability was then assessed by single-cell colony formation assay. (n = 4; 2 biologically independent ECFC donors, from 2 independent experiments; mean ± SD; p value was calculated by one-way ANOVA with multiple comparison)
Given these changes in gene expression, we next examined if SOX9 knockdown could affect EndMT at the functional level. Initial assessment through Matrigel capillary formation and single cell colony formation assay demonstrated a significant loss of endothelial-related functions after oxLDL treatment (p < 0.001 vs. ECFCshScr Veh, Fig. 4K-P). Unexpectedly, neither the capillary nor single cell colony formation capabilities of ECFCs were recovered by SOX9 silencing after oxLDL treatment. Indeed, the silencing of SOX9 alone in ECFCs in the absence of oxLDL exposure was sufficient in directly and significantly inhibiting ECFC self-renewal, with decrease in the percentage of HPP colonies formed comparable to oxLDL treated ECFCs (p < 0.001 vs. ECFC [9] Veh, Fig. 4Q). The low basal expression of SOX9 may therefore be needed to maintain the ECFC progenitor population. Upon shRNA silencing of SOX9, oxLDL did not further affect the endothelial function of ECFCs in terms of capillary formation (p = 0.0793) or HPP colony formation (p = 0.99) as compared to ECFC [9] exposed to vehicle alone.
To investigate the role of SOX9 on the gain of mesenchymal attributes upon oxLDL exposure, scratch mobility and 3D collagen gel contraction assays were used. As part of EndMT activation, endothelial cells display enhanced migratory functions, reflected here, with oxLDL treated ECFCshScr colonizing significantly higher percentage of scratched space compared to vehicle (p < 0.001 vs. ECFCshScr Veh, Fig. 4M and N). This increased mobility was abrogated within the ECFC [9] group. Furthermore, the gain of specific-mesenchymal function can also be demonstrated through the significant increase in cell contractility with more than 50% gel size reduction after 72 h (p < 0.001 vs. ECFCshScr oxLDL, Fig. 4O and P). This process is generally attributed to fibroblasts differentiating into a more contractile state as myofibroblast. This transition is also supported at the molecular level, as an array of myofibroblast specific genes (ACTA2, COL1A1, COL1A2, TGFB) were found to be significantly up regulated within the RNAseq after oxLDL treatment. Equally, this functional transition was efficiently reversed in ECFC [9] treated with oxLDL, demonstrating a central role of SOX9 in driving oxLDL-induced EndMT at both the functional and molecular levels. These findings corroborate that the silencing of SOX9 can effectively attenuate mesenchymal transition by inhibiting the aberrant expression of key drivers of this process (SNAI1, SLUG). However, it was not able to restore endothelial and progenitor function.
To further establish the importance of SOX9 in the EndMT phenotype, we next proceeded with its overexpression. ECFCs overexpressing SOX9 (OvSOX9) were compared to ECFCs harbouring only the empty vector (EV). As a result of the overexpression, SOX9 levels were more than 5000-fold increased in ECFC OvSOX9 (Supplemental Fig. 5A). This increased expression led to morphological alterations in the absence of oxLDL along with elevated expression levels of mesenchymal genes such as COL2A1 and SNAI1 or CD90 at the protein level (Supplemental Fig. 5B-4E). Functionally, overexpression of SOX9 dramatically reduced HPP colony forming potential resulting in increasing numbers of endothelial clusters pointing to a loss of self-renewal (Supplemental Fig. 5D). Similarly, SOX9 overexpression resulted in a reduction in tube formation (Supplemental Fig. 5F, G). Overall, SOX9 overexpression recapitulated gene expression and endothelial functional changes observed in EndMT induced by oxLDL.
Endothelial-specific ablation of Sox9 attenuates EndMT in vivoWe next examined if our observations in ECFCs could be reproduced in vivo. To examine this, Cdh5CreERt2/ROSAYfp/Sox9+/+ (Sox9eWT) and Cdh5CreERt2/ROSAYfp/Sox9flox/flox (Sox9eKO) mice were generated to examine the impact of endothelial-specific deletion of Sox9.
To assess the contribution of high fat diet (HFD) on endothelial SOX9 expression, and subsequent EndMT, mice were subjected to a 4-week HFD regime (Fig. 5A). At the end of the diet, significant increases in body weight and visceral fat mass within the HFD group was observed, regardless of genotype (p < 0.001 vs. Chow, E Supplemental Fig. 6A and B). Immunofluorescent examination of the coronary arterial vessels demonstrated a significant increase in nuclear expression of SOX9 in endothelial cells (CD31 + cells) of Sox9eWT mice fed a HFD compared to a normal chow diet, supportive of HFD inducing the expression of SOX9 (p = 0.0004 vs. Sox9eWT Chow, Fig. 5F and G). Next, we examined murine endovascular progenitors (EVPs) as a surrogate model of human ECFCs to evaluate progenitor activity [2,10,20,21,22].
Fig. 5
Endothelial-specific ablation of Sox9 attenuates high-fat diet-induced EndMT in vivo. (A) Schematic timeline and mouse model used throughout this experiment. Cdh5CreERt2 (Sox9eWT) and Sox9fl/fl/Cdh5CreErt2/ROSAYfp (Sox9eKO) were fed either the control chow or HFD. Aortic endothelial populations were then assessed 4 weeks after. (B) Gating strategy used to identify endothelial populations. (C-E) FACS analysis of Sox9eWT and Sox9eKO aortic endothelial populations after 4 weeks HFD. (F-G) Immunofluorescent image and quantification of endothelial SOX9 expression of Sox9eWT and Sox9eKO coronary arterial vessel after HFD. (H-I) Coronary arterial vessel co-expression of FSP-1, CD31 and Cre-YFP examined under immunofluorescence staining in Sox9eWT and Sox9eKO fed HFD. (n = 5; 5 biologically independent samples; mean ± SD; p value was calculated by one-way ANOVA with multiple comparison)
The aortic endothelial hierarchy was examined under flow cytometry as previously reported by us (as controlled by fluorescence-minus-one staining, Supplemental Fig. 6C). Briefly the YFP + LineagenegCD34+CD31low fraction identified endovascular progenitors (EVP) known to carry self-renewal activity. No significant differences in Cdh5Cre driven-YFP + cells (representing total endothelial cells) were detected between the Sox9eWT and Sox9eKO groups, regardless of diet. After gating on total YFP+Lineageneg endothelial cells, CD31 and CD34 were used to delineate the murine endothelial heterogeneity and hierarchy as previously reported [7,10,20]. In Sox9eWT mice fed HFD, a significant accumulation of mesenchymal M cells (YFP + LineagenegCD31-CD34-) indicative of EndMT (p < 0.002 vs. Sox9eWT Chow, Fig. 5B and E) was observed and was associated with a depletion of EVP progenitors (p < 0.001 vs. Sox9eWT Chow, Fig. 5B and C) corroborating the transition to a mesenchymal phenotype and loss of progenitor numbers observed in vivo. Upon Sox9 deletion, the transition of EVPs to M cells was effectively prevented in Sox9eKO group, as no significant changes were observed between the diet groups (Fig. 5B and E). However, EVP progenitor levels did not differ according to diet. Importantly, even in the absence of a high-fat diet, conditional loss of Sox9 in the endothelium resulted in a significant reduction in the progenitor population (p < 0.0024 Sox9eKO vs. Sox9eWT Chow), recapitulating the importance of the Sox9 gene in progenitor function and self-renewal.
Next, we used immunostaining to characterise cells labelled with YFP, witnessing their expression of the endothelial marker cdh5 at the time of tamoxifen injection. We used a well-characterised mesenchymal factor, fibroblast-specific protein 1 (FSP-1), in addition to a key endothelial marker CD31 to examine EndMT within the coronary vasculature. The number of YFP + cells which co-expressed CD31 and FSP-1 was significantly increased in Sox9eWT mice given HFD compared to chow supporting the advent of EndMT upon HFD (p < 0.001 vs. Sox9eWT Chow, Fig. 5H and I). Equally, in support of the previous flow cytometry observations, the conditional deletion of Sox9 within the endothelium largely attenuated HFD-induced EndMT, reflected by the reduction of endothelial FSP-1 expression.
Overall, these findings support our in vitro observation that HFD induces a mesenchymal transition associated with the loss of progenitors and the former can be inhibited effectively by the loss of Sox9.
留言 (0)