
記住我
We have previously shown that Nnat is crucial for insulin storage and GSIS in the mouse [35]. NNAT expression is diminished in rodent models of obesity and diabetes including the Zucker diabetic fatty (ZDF) rat model (85% decrease, p=0.0023, n=5 [45]) and obese ob/ob mice (ESM Fig. 1a). Pancreatic sections of db/db mice displayed a loss of NNAT immunoreactivity across islets, and a significant reduction of NNAT+ beta cells (ESM Fig. 1b,c).
Reanalysis of published scRNA-seq datasets from primary mouse beta cells at embryonic (embryonic day 17.5 [E17.5]) [39, 40] and adult [5, 41, 42] stages identified Nnat as amongst the most highly variable genes between individual cells (ESM Table 1). Pseudo-time analysis of embryonic (E12.5 to E17.5) mouse beta cells [39] clearly demarcated the differentiation of beta cells through embryonic cell states, with late embryonic (E17.5) beta cells overlapping with high levels of Nnat expression (Fig. 1a–c, ESM Figs 2a–f, 3a–k). Analysis of the proportions of different beta cell progenitors revealed that Nnat+ cells become more prominent at later stages of development (Fig. 1d). Integrating cells from the embryonic beta cell trajectory with adult beta cells [41, 42] allowed us to evaluate the progressive change in beta cell markers and Nnat expression from development to maturity (Fig. 1e,f, ESM Fig. 4a–i), and revealed large increases in Ins1 expression alongside Nnat downregulation in most beta cells. Nevertheless, a considerable number of adult beta cells (with high expression of Ins1/Ins2) also expressed Nnat (Fig. 1f). Thus, Nnat appears to mark late-stage beta cell differentiation, peaking in expression around E17.5 and then gradually being downregulated across most beta cells in adulthood.
Fig. 1
Nnat expression is a marker for late-stage beta cell differentiation during islet development. (a–c) UMAP projection of scRNA-seq data from embryonic mouse islets (E12.5–E17.5), with cells labelled by their cellular state (a), pseudo-time (scale bar represents abstract unit of time) (b) and corresponding Nnat expression (log-normalised expression level) (c). (d) Changes in the distribution of beta cell precursors from E12.5 to E17.5. (e, f) Violin plots showing the fluctuations in Ins1 (e) and Nnat (f) expression (both log-normalised expression level) from E12.5 to adulthood in the beta cell development trajectory. Prolif., proliferating; UMAP, uniform manifold approximation and projection
In light of the abundance of Nnat+ beta cells during late embryogenesis, we further evaluated the E17.5 dataset [39] and again identified two beta cell populations with distinct levels of Nnat expression (Fig. 2a–c, ESM Fig. 5a–f). Embryonic Nnat+ beta cells were enriched for Ins1, Ins2, Nkx6.1, Pdx1, Ucn3, Slc2a2, Iapp, Ero1b, G6pc2, Dlk1 and Npy expression compared with Nnat− beta cells which had higher Neurog3, Pax4, Gcg, Arx and Ghrl expression, indicating that the Nnat+ beta cells were more fully differentiated (Fig. 2d, ESM Table 2). Nnat+ beta cells were also de-enriched for Cd24a (Fig. 2d, ESM Table 2). Gene set enrichment analysis (GSEA) revealed upregulation of processes related to protein synthesis, transport from endoplasmic reticulum (ER), ER stress, as well as oxidative phosphorylation and carbohydrate metabolism, with downregulated processes including mRNA processing, splicing and histone methylation (Fig. 2e). Indeed, a separate cell clustering analysis [46] identified Nnat as a highly differentially expressed gene between two beta cell clusters at both the late embryonic [39, 40] and adult [5] stages (ESM Figs 6a–h, 7a–g, ESM Table 3).
Fig. 2
Transcriptomic analyses of islet cells at E17.5 reveal Nnat+ cells to be a more differentiated beta cell subcluster. (a, b) Islet cells plotted in the UMAP space and coloured by Ins1 (a) and Nnat (b) expression, respectively. Individual cellular clusters are labelled. Beta cell clusters are highlighted with black circles. (c) Distribution of Nnat expression among beta cell clusters, along with the expression range used in defining Nnat+ beta cells (highlighted by the red rectangle). (d) Volcano plot showing the log2 fold change in gene expression between Nnat+ and Nnat− beta cells against FDR-corrected p values. Genes with FDR<0.1 are coloured red. Top ten upregulated and downregulated genes are labelled by gene name. (e) GSEA of differentially expressed genes between Nnat+ and Nnat− beta cells (using gene ontology-biological process [GO-BP] terms), showing the top ten upregulated and downregulated processes. The normalised enrichment score (NES) is plotted along the x-axis. Circle diameters are proportional to the number of genes in each process, with the colours defining statistical significance. UMAP, uniform manifold approximation and projection
We confirmed some of these findings at the protein level using immunofluorescence in postnatal and adult mice (Fig. 3a,b). We noted that NNAT protein expression is highly dynamic in beta cells across the mouse islet throughout the postnatal stage, transitioning from expression throughout the majority (90.4±3.2%, n=15 mice) of beta cells at late embryogenesis to a subset (24.4±3.0%, n=5 mice) of beta cells by postnatal day 14 (P14) (Fig. 3a,b). NNAT+ beta cells persisted throughout the islet into adulthood (Fig. 3a,b). NNAT expression was not apparent in other mouse islet cell types, including alpha and delta cells, at P14. However, NNAT expression was detectable in a small number of alpha and delta cells at E17.5 (Fig. 3c).
Fig. 3
A subpopulation of NNAT+ beta cells develops during the early postnatal period in mice. (a) Representative confocal microscopy of pancreatic cryosections from wild-type mice on a C57BL/6J background of developmental stages from E17.5 through the postnatal period into adulthood. Sections were immunostained with antibodies against endogenous NNAT (green) and insulin (INS, red). Nuclei are visualised with DAPI and sections from P56 mice with constituent deletion of Nnat were used as an immunostaining control. Scale bar, 100 μm (inset, 10 μm). (b) Quantification of NNAT+ beta cells from images shown in (a), expressed as NNAT/INS co-positive cells as a percentage of total INS-positive cells (n=4–15 mice per timepoint, Kruskal–Wallis test with Dunn’s multiple comparisons). (c) Representative confocal microscopy of pancreatic cryosections as in (a) from E17.5 and P14 mice (n=18 and n=5 mice, respectively) immunostained with antibodies against endogenous NNAT (green) and GCG or SST (both grey). Scale bar, 100 μm. Representative images from three independent experiments and breeding pairs. *p<0.05, **p<0.01, ***p<0.001. GCG, glucagon; KO, knockout; SST, somatostatin
NNAT immunoreactivity is detected in a subset of beta cells in human islets and NNAT deficiency in human beta cells blunts GSISWhile the role of NNAT has previously been examined in mouse beta cells [35], no data currently exist for human beta cells. We therefore next compared the functionality of human NNAT+ and NNAT− beta cells. Transient silencing of NNAT in human EndoC-βH1 beta cells completely abrogated glucose-stimulated insulin release (Fig. 4a–c). As in mouse beta cells [35], NNAT interacted in cellulo with subunits of the SPC, SEC11A, SPCS1, SPCS2 and SPCS3 (ESM Fig. 8a), likely via SPCS1 (ESM Fig. 8b).
Fig. 4
NNAT is expressed in a subset of human beta cells and NNAT deficiency in human beta cells reduces insulin secretion. (a) GSIS analysis at low (3 mmol/l) and high (16.7 mmol/l) glucose in human EndoC-βH1 beta cells following 72 h of lentiviral-mediated shRNA knockdown of NNAT with data expressed as insulin secreted into culture media as a percentage of total cellular insulin content (n=8 independent cultures per group, two-way ANOVA with Sidak’s multiple corrections test). (b, c) RT-PCR (n=8, unpaired Student’s t test) and western blot (n=4, Mann–Whitney U test) analysis of NNAT expression in EndoC-βH1 beta cells after transient NNAT silencing as in (a). (c) Western blotting analysis of NNAT protein levels shown via a representative blot of two independent experiments. β-Tubulin was used as a loading control. Mean values for band intensities in multiple experiments quantified by densitometry are shown below the panel, expressed relative to scramble (SCR) shRNA controls. (d) Representative confocal microscopy of human pancreatic cryosections from younger (15.6±0.9 years, n=5) and older (71.0±3.9 years, n=4) donors. Sections were immunostained with antibodies against endogenous NNAT (green), insulin (INS, red) and glucagon (GCG, grey). Donor sex and age are indicated in the text. Nuclei are visualised with DAPI. Scale bar, 100 μm. (e, f) Quantification of NNAT+ beta (e) and alpha (f) cells from images shown in (d), expressed as NNAT/INS or NNAT/GCG co-positive cells as a percentage of total INS-positive or GCG-positive cells. *p<0.05, ***p<0.001
Heterogeneous expression of NNAT in human islet cells was also observed in whole pancreatic sections from multiple donors. Thus, NNAT was expressed in a subset of beta cells in both younger (15.6±0.9 years, 15.0±4.0%, n=5) and older (71.0±3.9 years, 17.7±2.1%, n= 4) donors (Fig. 4d,e, ESM Fig. 9a, ESM Table 4, ESM Human Islet Checklist). Interestingly, NNAT was also expressed in a large fraction of human alpha cells (but not in delta cells, Fig. 4d,f, ESM Fig. 9b) in both younger (47.0±2.2%) and older donors (69.6±12.2%).
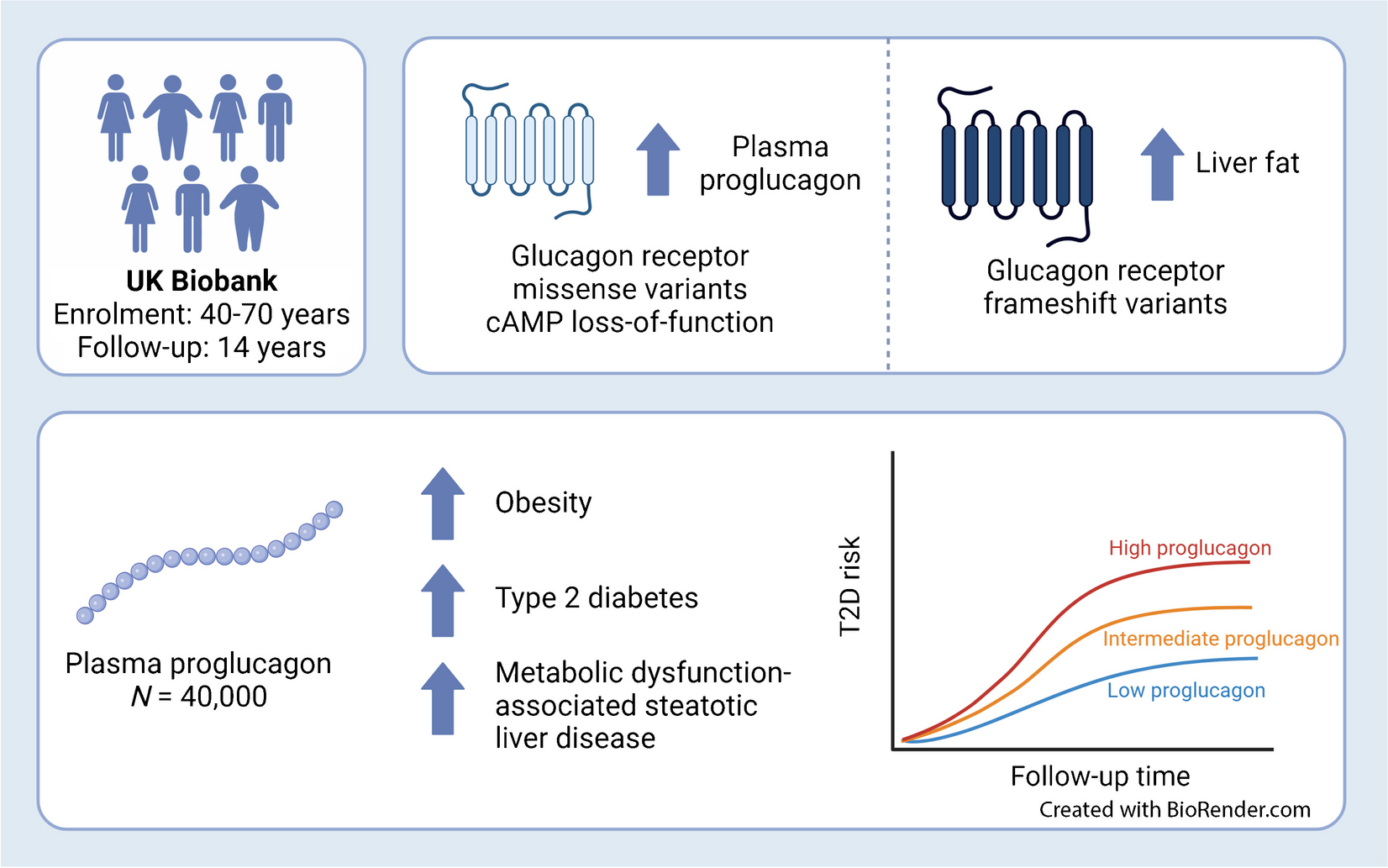
Heterogeneous NNAT expression in beta cells during postnatal development is associated with altered CpG methylationTo extend the findings above using an orthogonal approach, and to facilitate subsequent functional studies, we utilised a Bacterial Artificial Chromosome (BAC) reporter mouse line expressing eGFP under the control of the Nnat promoter and upstream enhancers (ESM Results, ESM Fig. 10a–e). Aligning with studies of the endogenous gene (Figs 1, 2, 3 and 4), adult reporter mice expressed eGFP in a subpopulation (~15% of total) of beta cells (Fig. 5a).
Fig. 5
Beta cell heterogeneity of NNAT expression is associated with changes in CpG methylation at the Nnat promoter. (a) Representative confocal microscopy of pancreatic cryosections from P56 mice with Nnat-driven eGFP expression from the paternal allele (NnatWT/eGFPpat) (n=7 mice). Sections were immunostained with antibodies against eGFP (green) and insulin (INS, grey). Nuclei are visualised with DAPI. Scale bar, 50 μm. (b) FACS separation of dispersed primary islet cells from reporter mice with insulin-driven expression of tdTomato (to label beta cells) and Nnat-driven eGFP expression from the paternal (NnatWT/eGFPpat) or maternal (NnatWT/eGFPmat) allele or wild-type (NnatWT/WT) at this locus (representative FACS plot of the dispersed islet preparation from a single mouse shown) (n=8, 3 and 3 mice per genotype, respectively, Kruskal–Wallis test with Dunn’s multiple comparisons). (c) Quantification of data in (b) expressed as percentage of eGFP/tdTomato co-positive primary islet cells. (d) Representative bisulphite analysis of CpG methylation at the Nnat promoter in FACS-purified islet cell populations from (b) (n=3 NnatWT/eGFPpat mice with paternally expressed Nnat-driven eGFP, n≥12 clones each). Closed circles, methylated CpG; open circles, unmethylated CpG. (e, f) Quantification of data in (d) expressed as percentage CpG methylation across the Nnat promoter at individual CpGs (e) and across the entire Nnat promoter (f) (both paired Student’s t test). (g) Schematic summarising level of CpG methylation at the Nnat promoter and gametic DMR in NNAT+ vs NNAT− beta cells (created with BioRender.com). Representative image in (a) and bisulphite analyses in (d) used experimental mice from three independent experiments and breeding pairs. *p<0.05, **p<0.01
To isolate and purify beta cells based on NNAT levels, we crossed Nnat-eGFP reporter mice to animals expressing Cre recombinase under the RIP [43] and to transgenic mice expressing a tdTomato reporter downstream of a loxP-flanked stop codon [44]. Dispersion of primary adult islets into single cells, and subsequent FACS analysis, verified the presence of NNAT+ and NNAT− beta cells, with the NNAT+ fraction composing 15.3±1.8% (n=8 mice) of the total beta cell compartment (Fig. 5b,c, ESM Figs 11a,b, 12a–f). Collection of FACS-purified NNAT+ and NNAT− beta cells and subsequent bisulphite sequencing analysis revealed that CpG methylation at the gametic DMR (known to control monoallelic Nnat expression; Introduction) was unchanged (ESM Fig. 11c–e). Nevertheless, CpG methylation at the Nnat promoter was significantly altered across this genomic region, with minimal CpG methylation observed in NNAT+ beta cells (Fig. 5d–f).
Recent whole-genome methylation analysis in mouse germ cells has revealed that the Nnat promoter region is unmethylated in sperm, and this was confirmed by targeted bisulphite sequencing analysis (ESM Fig. 11f–h), whereas it is fully methylated in oocytes [47]. Thus, the ‘classical’ imprinting of Nnat is unaltered between beta cell subtypes with near binary (‘on/off’) expression of NNAT. However, and overlaying this control, a second DMR at the Nnat promoter within the non-imprinted allele dictates beta cell subtype specificity of expression (Fig. 5g).
The de novo DNA methyltransferase DNMT3A establishes NNAT beta cell subtype specificityTo ask whether this apparent transition in Nnat expression in the early postnatal period is driven by de novo CpG methylation, we assessed endogenous NNAT beta cell immunoreactivity in postnatal mice conditionally deleted for the methyltransferase DNA methyltransferase 3 alpha (DNMT3A) at the pancreatic progenitor stage (using Pdx1-Cre) [6]. NNAT staining in control mice at P6 demonstrated expression in a subpopulation of beta cells, whereas deletion of DNMT3A resulted in a loss of this heterogenous expression across the islet (Fig. 6a,b, ESM Fig. 13a). These findings demonstrate that de novo methylation is likely to drive NNAT restriction across the beta cell complement in the first few days of postnatal life.
Fig. 6
Postnatal restriction of NNAT in a subset of beta cells is at least partially driven by the de novo methyltransferase DNMT3A. (a) Representative confocal microscopy of pancreatic cryosections from mice with conditional deletion of DNMT3A under the control of the Pdx1 promoter (Pdx1-Cre+ Dnmt3afl/fl) vs control (Pdx1-Cre− Dnmt3afl/fl) mice at P6. Sections were immunostained with antibodies against endogenous NNAT (green) and insulin (INS, red). (b) Quantification of NNAT+ beta cells from images shown in (a), expressed as NNAT/INS co-positive cells as a percentage of total INS-positive cells. Scale bar, 50 μm (n=8 mice per genotype, unpaired Student’s t test, **p<0.01). Nuclei are visualised with DAPI
NNAT+ and NNAT− beta cells have distinct transcriptional signaturesIn contrast to the embryonic populations described above, RNA-seq revealed that FACS-sorted adult (8 weeks) NNAT+ and NNAT− beta cells displayed high transcriptional overlap and were similar to each other when compared with non-beta endocrine cells (Fig. 7a). Major beta cell identity markers such as Ins1, Ins2, Mafa, Slc2a2 (Glut2) and Nkx6.1 were not differentially expressed (ESM Table 5).
Fig. 7
NNAT+ adult beta cells are transcriptionally distinct and have significantly higher insulin content. (a) Correlation matrix of differentially expressed genes between NNAT+ and NNAT− beta cells as assessed by RNA-seq analysis (n=4 FACS-purified populations from individual mouse islet preparations). Scale bar (0–255) gives an integer provided to the R function where the darker colour corresponds to higher sample correlation. (b) Heatmap of the top ten most differentially expressed genes reduced in NNAT+ beta cells compared with NNAT− beta cells. (c, d) GSEA showing categories significantly enriched (c) and reduced (d) in NNAT+ (vs NNAT−) beta cells. (e, f) Representative confocal microscopy of pancreatic cryosections from P56 (8-week-old) wild-type mice on a C57BL/6J background immunostained with antibodies against endogenous NNAT (green) and UCN3 (red, e) or TOM20 (red, f). Scale bar, 100 μm. Nuclei are visualised with DAPI. Representative images from three independent experiments and breeding pairs (g, h). Quantification of NNAT (g) and TOM20 (h) staining in NNAT− and NNAT+ beta cells (INS+) from images shown in (f), expressed as mean intensity of the NNAT or TOM20 channel (*p<0.05, paired Student’s t test, 1329 NNAT− and 295 NNAT+ beta cells from 15 islets, n=3 mice). (i) Insulin content assessed in NNAT+ and NNAT− beta cells (*p<0.05, n=7 FACS-purified populations from individual mouse islet preparations, Wilcoxon matched-pairs signed rank test). AU, arbitrary units; FC, fold change
Nevertheless, we identified 241 (1.8%) and 79 (0.6%) genes that were significantly down- and upregulated in NNAT+ vs NNAT− beta cells, respectively (>2-fold, false discovery rate [FDR]<0.01). Differentially expressed genes included several markers of non-beta cell islet lineages (Pyy, Ppy, Mafb, Sst) that were lower in the NNAT+ beta cell fraction, genes linked with beta cell heterogeneity and plasticity (Gpx3, Rbp4), the beta cell immaturity marker Cd81 [20] and the Cd24a antigen [21] (Fig. 7b). Interestingly, Npy was enriched in NNAT+ beta cells (ESM Table 5), consistent with our observations at late embryogenesis (E17.5, Fig. 2d). We did not observe any clear differences between NNAT+ and NNAT− beta cells in the expression of other imprinted genes known to be functional in the beta cell (Plagl1/ZAC, Dlk1, Rasgrf1, Cdkn1c, Grb10 and Gtl2/MEG3), nor changes in ‘disallowed’ genes whose expression is known to be restricted in mature functional beta cells (Hk1, Mct1 [Slc16a1] and Ldha) [48] (ESM Table 5). Likewise, no differences were apparent in the levels of transcripts encoding proteins previously demonstrated by others to mark specific beta cell subpopulations, such as Flattop/Cfap126 [16], CD9 and ST8SIA1 [10], Cd63 [23], ‘virgin’ beta cells (via Ucn3) [24] or those differentially expressed in beta cells implicated in the control of Ca2+ dynamics (‘hubs’ [14], and ‘leaders’ [15, 26]).
GSEA did, however, reveal enrichment of pathways in NNAT+ beta cells including translation initiation, the electron transport chain (ETC), oxidative phosphorylation and signal recognition particle (SRP)-dependent co-translational protein targeting (Fig. 7c). Pathways involving genes associated with hepatocyte nuclear factor 1 alpha (HNF1A) and GLI-similar family zinc finger 3 (GLIS3) targets were reduced in the NNAT+ beta cell fraction (Fig. 7d). NNAT+ beta cells displayed higher levels of the late maturation marker urocortin 3 (UCN3) (Fig. 7e, 0.357±0.009 in NNAT+ vs 0.264±0.004 arbitrary units in NNAT−, **p<0.01, 290 NNAT+/1876 NNAT− beta cells from 13 islets/n=3 mice) and a small but significant increase in translocase of outer mitochondrial membrane 20 (TOM20) immunostaining, consistent with a higher mitochondrial volume or number (Fig. 7f–h). Furthermore, FACS-sorted primary NNAT+ beta cells had a significantly higher insulin content than NNAT− cells (Fig. 7i).
The NNAT+ beta cell population shows impaired glucose-stimulated Ca2+ dynamics and is de-enriched for highly connected ‘hub’ cellsTo determine whether NNAT may influence beta cell connectivity and membership of the ‘hub’ cell subgroup [14], we studied glucose-induced Ca2+ dynamics in Nnat-deficient (Nnat+/−p) islets [35]. Ca2+ increases in response to high (11 mmol/l) glucose were significantly higher in Nnat+/−p than control islets, and while the mean Pearson’s coefficient of correlation and proportion of highly connected ‘hub’ cells were not significantly different between Nnat-deficient and control islets (ESM Results, ESM Fig. 14a–d), wild-type ‘hub’ cells were significantly more connected than Nnat-deficient ‘hub’ cells (ESM Fig. 14e,f). Thus, NNAT is a marker of less well-connected cells.
We further explored this question using islets from Nnat-eGFP reporter mice, using the red-shifted calcium probe Cal-590 AM [49]. Nnat-GFP+ and Nnat-GFP− cells responded similarly to challenge with high (11 mmol/l) glucose (Fig. 8a). Beta cell–beta cell connectivity (Fig. 8b) was not different between NNAT+ and NNAT− populations and, considered across the whole population, the mean Pearson’s coefficient of correlation was 0.87±0.01 (Fig. 8c).
Fig. 8
NNAT+ beta cells show altered glucose-induced Ca2+ dynamics and are de-enriched for highly connected ‘hub’ cells within individual islets. (a) Ca2+-bound Cal-590 AM fluorescence in response to high glucose (11 mmol/l) in NNAT− and NNAT+ cells from primary islets from NnatWT/eGFPpat reporter mice expressed as normalised intensity over time (F/Fmin) (n=61 islets total from five mice per genotype; quantification of AUC on the right, p=0.063, Wilcoxon matched-pairs signed rank test). (b) Representative Cartesian map of beta cells with colour-coded lines connecting cells according to the strength of coactivation (colour-coded R values from 0 to 1, blue to red). Beta cells are represented by differently coloured nodes depending on their coactivity with the other beta cells, where black nodes indicate coactivity with ≥80% of the remaining beta cells, while grey, white and orange nodes represent coactivity with ≥60%, ≥40% and <40%, respectively. Nodes circled with a solid black line indicate NNAT+ cells. (c) Representative heatmaps depicting connectivity strength (r) of all cell pairs (colour-coded r values from 0 to 1, blue to yellow). (d) Log–log graphs of beta cell–beta cell connectivity distribution. NNAT+ cells are represented by green circles while NNAT− cells are represented by red circles (45 islets total using primary islet preparations, each from an individual NnatWT/eGFPpat reporter mouse). (e) Categorisation of beta cells based on data from (d). (f) Percentage coactivity of beta cells between all cells in identified ‘hub’ and ‘follower’ cells. (g) The proportion of cells designated as ‘hub’ vs ‘follower’ cells in both the NNAT− and NNAT+ populations assessed in each of 45 islets (**p<0.01, Wilcoxon matched-pairs signed rank test). Analyses in panels (a–g) obtained from three independent experiments and three different breeding pairs of experimental mice. AU, arbitrary units
Of all cells examined, 4.27% were NNAT− ‘hub’ cells (Ca2+ responses with at least 30% of all cells, Fig. 8d) connected to an mean of 39.0% of all beta cells (Fig. 8e,f). The fraction of these (1.15%, Fig. 8e) identi
留言 (0)