

記住我
Migraine and associated symptoms are a significant burden to the financial, mental, and physical wellbeing of those that live with this chronic disease. A retrospective study conducted in the United States showed that migraine patients spent as much as $11,010 annually in direct healthcare costs on average in an attempt to treat their disease which included frequent emergency room, inpatient and outpatient visits and medication prescriptions (Bonafede et al., 2018). In the same study, the annual indirect costs of migraine patients due to disability and workplace absenteeism were about $2350 higher as compared to matched non-migraine patients. In a survey conducted on about 160,000 Americans aged 12 years and above in 2012 under the American Migraine Prevalence and Prevention Study (AMPP), and in line with other studies, the prevalence of migraine was higher in females compared to males across all age groups. A total 7.68% of the study population had migraine and within this group 0.91% were chronic migraine patients (Buse et al., 2012). Chronic migraine was also associated with the highest headache related disability. It is important to note the observation that within this study chronic migraine prevalence was higher in low-income households. Although it has been reported that high income developed countries have statistically higher migraine populations than poorer countries, this may be due to the point that more studies are conducted in high income countries which could skew data (Stovner et al., 2022). Migraine has a sex-related pattern as well and with a global prevalence of 11.24 and 19.01%, respectively, for males and females using data obtained from 204 countries and territories in 2019 (GBD 2019; Diseases and Injuries Collaborators, 2020). A clinically observed sex-related difference is important to consider in drug discovery research due to the implications related to chronic migraine preclinical model validation. Despite significant financial, mental and physical burdens, a need persists for a better understanding of chronic migraine neurobiology, underlying receptor mechanisms, as well as therapeutic treatment options.
The understanding of migraine pathophysiology has evolved over the years. There are several different theories surrounding migraine development and maintenance. These theories include: the vascular and the neurovascular theories of migraine (Graham, 1938; Goadsby et al., 1990). Each of these theories will be discussed in this review. One of the earliest theories postulated was the vascular theory. A study observed that ergotamine tartrate, through its vasoconstrictive effects, was able to reduce the pulsations as well as headache associated with migraine (Graham, 1938). The importance of vasodilation in migraine pathophysiology has been further investigated by observing changes that occur in blood vessels within the brain. It was initially hypothesized that dilation of the temporal and middle meningeal arteries might play a role (Graham, 1938). Although it was observed that migraine relief could be produced after compressing the temporal artery, it was shown to occur in only few migraine patients (Drummond and Lance, 1983). The external carotid artery has also been implicated to be involved in migraine headache (Ray and Wolff, 1940). The vascular theory has remained a controversy due to emerging evidence that refutes this explanation, including the finding that not all vasodilators cause migraine. In a randomized clinical trial that involved the infusion of vasoactive intestinal peptide (VIP) into 12 patients, migraine-like headache was not triggered in migraine patients, although there was vasodilation of the superficial temporal artery (Rahmann et al., 2008). In another study that involved the use of adrenomedullin to induce migraine headache, despite a vasodilatory response seen in blood vessels, there was no difference compared to placebo (Petersen et al., 2009) and these findings have been replicated in a preclinical study involving a mouse migraine model (De Logu et al., 2019). However, it is still recognized that vasodilation is an important mechanism that contributes to migraine and some current human and animal models of migraine still rely on vasodilatory properties of migraine headache inducers to assess novel compounds. Critical unknown migraine pathophysiological areas that are currently of interest in the migraine research community include: determining an initial trigger for migraine, the role of central vs. peripheral migraine headache mediation, the contributing factors that lead to either an acute or sustained migraine attack, as well as biomarkers that correlate with migraine symptoms.
Currently, the prevailing theory embraces the role of inflammation and neural release of peptides in the development of migraine headache in addition to the vascular theory, and migraine is thus considered to be a neurovascular condition. This review will provide a cursory overview of the dominant processes that contribute to migraine pathophysiology. It will also cover the associated clinical features of migraine, human and animal models of migraine with their strengths and weaknesses, migraine therapeutics as well as areas that could be considered for future research. Moreover, a major emphasis will be placed on preclinical migraine models and experimental therapeutic development of antimigraine drugs. Although important to migraine and associated symptomology, this review will not cover in vitro models of migraine, other migraine types including acute and chronic migraine with or without aura, non-pharmacological treatments, and in-depth mechanisms of action for the different treatment options. For further review please see the following references on pathophysiology of migraine (Goadsby et al., 2017), experimental models of migraine (Harriott et al., 2019), and treatment options for migraine (Zobdeh et al., 2021).
The literature search was conducted using PubMed, Web of Science and SCOPUS using the search words: migraine, cortical spread depression, migraine pathophysiology, and migraine treatment. The definitions of migraine are limited to that provided by the International Headache Society (Headache Classification Committee of the International Headache Society (IHS), 2018). In databases that allowed filtering search results, the search was limited to original research articles. Original research articles were screened to identify experimental work carried out on rodents while clinical research papers were limited to randomized clinical trials and meta-analysis. All research included in this review needed to fulfill the requirement of appropriate treatment and control groups with objective and unbiased conclusions. Research papers that utilized non-pharmacological interventions were excluded.
1.1 What is migraine?In discussing migraine, it is imperative to distinguish an acute migraine from other headache classes, as well as chronic migraine from chronic headaches. Headache is a subjective symptom which is commonly reported among patients and ranges in severity from mild to severe. Its impact on health is so profound that it ranked in 2019 according to the Global Burden of Disease Study among the top ten conditions for disability-adjusted life-years (DALYs). DALYs are a measure of the number of years lost due to poor health, disability or early death to express disease burden (GBD 2019; Diseases and Injuries Collaborators, 2020). To facilitate diagnosing criteria for different types of headache, the International Headache Society (IHS) is responsible for publishing the International Classification of Headache Disorders and their third edition was published in 2018 based on emerging scientific evidence on headaches (Headache Classification Committee of the International Headache Society (IHS), 2018). Primary headaches are headaches without an underlying cause, and migraine in addition to tension-type headache are the two most common primary headache disorders with a quantifiable impact on global population health (Steiner and Stovner, 2023). It is also worth noting that migraines have been shown to be the second highest cause of disability among all populations globally using age-standardized years of life lived with disability (YLD) as a metric (Stovner et al., 2018; Steiner and Stovner, 2023). The loss of productivity associated with migraines is evident since the group most affected form the largest proportion of the workforce that drives national economies. The migraine disability assessment (MIDAS) questionnaire is a 5-point validated questionnaire that is also used to grade migraine associated disability using a scoring system (Stewart et al., 2000, 2001). According to the MIDAS tool, disability associated with migraine is described as the number of days missed at work or school, reduced productivity at home, work, and school as well as reduced social activities within 3 months (Stewart et al., 2000, 2001). After summing the number of days associated with headache/migraine-related disability, the total obtained is used to categorize severity into grades as shown in Table 1.
Acute migraine is described as headache that occurs unilaterally with a pulsating sensation, which lasts between 4 to 72 h and is associated with nausea and/or light sensitivity (photophobia) and sound sensitivity (phonophobia) (Headache Classification Committee of the International Headache Society (IHS), 2018). This headache class may be of moderate to severe intensity and may be exacerbated by physical activity. In some instances, the headache may be described as pressure, stabbing or aching (Kelman and Tanis, 2006). A patient who experiences headache, but not necessarily a migraine, on 15 or more days within a month for more than 3 months, with features of migraine headache on at least 8 days per month, can be said to have chronic migraine based on the ICHD-3 (Headache Classification Committee of the International Headache Society (IHS), 2018). Importantly, a challenge that these diagnostic criteria poses is the reliance on the patient’s ability to recall number of headache days accurately. It has been argued that the current definition of chronic migraine be reconsidered, and the threshold of days lowered since there are patients that have about 8 to 14 headache days per month but with a similar disease burden as 15 days or above (Ishii et al., 2021). These patients that experience a lower threshold may fail to get the needed medical treatment and represents another challenge in the treatment of chronic migraine.
1.2 Migraine with and without auraAccording to the studies, there are two types of migraine classified based on the presence or absence of aura, i.e., migraine with aura and migraine without aura (Rasmussen and Olesen, 1992; Headache Classification Committee of the International Headache Society (IHS), 2018). An aura describes reversible neurological symptoms that have a short duration and may manifest before a migraine headache or occur concurrently with the headache. These may include one or more of the following symptoms: visual (an absolute or partial blind spot at a fixed point with zigzag pattern, which has a distorted and bright edge of convex shape), sensory disturbances (numbness, migratory pins and needles sensation on body, face and/or tongue), speech disturbance (aphasia characterized by difficulty in expressing oneself), motor weakness, brainstem symptoms (tinnitus, vertigo/dizziness, diplopia/double vision, dysarthria/difficulty speaking, decreased consciousness).
Other classifications of migraine exist which are based on their unique clinical presentations which may differ in onset, frequency, or associated symptoms. For instance, hemiplegic migraine describes migraine with associated motor weakness, while migrainous infarction is a type of migraine that co-occurs with ischemic infarctions in the brain. A detailed list of these classifications is given by the International Headache Society in the International Classification of Headaches, 3rd Edition and are out of the scope of this review.
1.3 Symptoms associated with migraineThere are some patients who experience a prodromal phase (also referred to as premonitory phase) that heralds an impending migraine headache (Giffin et al., 2003) and may start hours or a few days to the attack; it may manifest as tiredness, photophobia and/or phonophobia, or nausea. Yawning was shown to be a common prodromal symptom in a cross-sectional study of 2,714 patients conducted by Laurell et al. (2016). It must be noted that this prodromal phase is different from an aura since it does not involve neurological symptoms. A post-dromal phase has also been described and occurs following headache resolution. But it can last as long as 48 h and some of its symptoms include fatigue, difficulty in concentration and stiffness of the neck (Headache Classification Committee of the International Headache Society (IHS), 2018). In a study of 11,388 migraine patients, cutaneous allodynia was reported among about 63% of patients (Lipton et al., 2008). Cutaneous allodynia tends to be frequently associated with migraine and may occur with or without migraine headaches and some unilateral autonomic symptoms such as nasal congestion, lacrimation and eyelid edema which are associated with trigeminal autonomic cephalalgias have also been reported among migraine patients (Barbanti et al., 2002). Other symptoms reported include diarrhea, taste abnormality, osmophobia (Kelman and Tanis, 2006).
1.4 Migraine risk factors 1.4.1 Obesity and depressionObesity has been shown to be a risk factor for the development of migraine (Santos et al., 2015; Gelaye et al., 2017; Miri et al., 2018) and it is hypothesized that increased levels of inflammatory mediators, increased sympathetic activity, and increased leptin (Pisanu et al., 2017) which are associated with obesity might influence the development of migraine. Adiponectin is also suspected to play a role (Duarte et al., 2014; Abbasi et al., 2019; Joshi et al., 2020) although there are contrasting opinions on this (Dearborn et al., 2014; Peterlin et al., 2016). Depression is another risk factor for migraine development (Breslau et al., 2003; Karakurum et al., 2004) as well as episodic to chronic migraine progression. An episodic migraine is migraine that does not occur as frequently as occurs in chronic migraine i.e., less than 15 days per month. Results from the observational cohort study of the AMPP study showed patients who experienced episodic migraine with depression were at a higher risk of chronic migraine onset and this could be explained by genetic predispositions or environmental factors such as life events or stress that have an influence on both conditions (Ashina et al., 2012).
1.4.2 Miscellaneous risk factorsOne unexplained risk factor is genetic susceptibility to raised serum calcium levels. Results obtained in a Mendelian randomization study showed hereditary patterns in both migraine development as well as hypercalcemia, although the underlying mechanism that links these two conditions is unknown (Yin et al., 2017). Calcium channel blockers such as flunarizine, a non-selective calcium channel blocker that are used to prevent migraine headaches give some level of credence to the involvement of calcium ions in migraine (Li et al., 2011). Also, asthma is suspected to be a risk factor that leads to the development of chronic migraine in patients with pre-existing episodic migraine. In a study based on results of the AMPP, it was found that episodic migraine patients with migraine were twice as likely to progress to chronic migraine compared to those without asthma (Buse et al., 2012; Martin et al., 2016). It is suspected that the upregulation of inflammatory mediators in asthma may play a role in this.
1.5 Migraine triggersMigraine patients have self-reported factors that trigger attacks. In a survey conducted on patients with migraine, triggers identified were classified into precipitating and aggravating factors. Some precipitating triggers which were predominant among migraine patients included weather changes, smell, smoke and light, while aggravating factors also included physical activity, noise and motion that involved straining and bending over (Spierings et al., 2001). Other triggers include stress and fatigue, lack of sleep, not eating on time, menstruation, and this has been corroborated in other studies (Ierusalimschy and Moreira Filho, 2002; Andress-Rothrock et al., 2010; Hauge et al., 2010). Recognizing the predominance of migraines in females and menstruation as a trigger, ICHD-3 has a category for menstrual migraine which is differentiated based on whether migraines occur during menstruation only, during and outside the menstruation period, or outside the menstruation period. Some foods are also an identified trigger for migraine although a mechanism remains elusive. A survey conducted in an outpatient’s neurology clinic in Turkey on 146 migraine patients showed that migraine headache was triggered by intake of chocolate, coffee, alcohol, milk and cheese although this was in a small percentage of participants (Hauge et al., 2010). In a randomized controlled trial among 38,370 females that sought to quantify the triggering of migraine on consuming some foods such as alcohol, ice cream, citrus fruits, milk and processed meat, it was found that dairy products and chocolate could be linked to the onset of migraine with aura (Rist et al., 2015). This study relied upon the consumption of low food quantities as being a marker of the food being a potential trigger for migraine, which was a limitation. It could be that the low intake of these foods was due to other factors such as general dislike for the food, and there is no evidence to suggest that food intake contributed to the migraine headache experienced.
2 Pathophysiology, pharmacological targets and United States Food and Drug Administration (US FDA) approved drugs for migraine 2.1 Pathophysiology and pharmacological targets 2.1.1 Cortical spreading depression (CSD) as a trigger of migraine auraAn attempt to understand the underlying mechanisms that cause migraine aura led to the discovery of a phenomenon known as cortical spreading depression (CSD). It has been described as a slow spreading wave of depolarization in glial and neuronal cells and is seen as a slow direct current potential shift due to high extracellular potassium ions with high intracellular sodium and calcium ions (Leao, 1947). CSD has been observed in migraine with aura where the CSD threshold is lowered to facilitate triggering an aura and seemly is the pathway through which some treatments act (Ayata et al., 2006). For example, some migraine prophylactic drugs were observed to decrease chemically-induced CSD in rats with prolonged administration having a sustained negative effect on CSD (Ayata et al., 2006). One of its downstream events is trigeminovascular system activation, which facilitates the release of nitric oxide to cause vasodilation, leading to increased blood flow in the meningeal artery. In addition, neurogenic inflammatory substance release in the dura mater causes sensitization of meningeal nociceptors leading to migraine headache. Experimentally, in rats, the onset of CSD produced Fos release in the trigeminal nucleus caudalis and downstream second-order neuronal activation, which led to migraine-like vascular events (Bolay et al., 2002). This phenomenon plays such an integral role in migraine that it is also used as a tool to monitor the efficacy as well as pharmacological activity of anti-migraine drugs (Schain et al., 2019). Although CSD is a spontaneous event, it occurs under the influence of preliminary events in the brain including neuronal hyperexcitability. While CSD is linked with the initiation of migraine with aura, it does not seem to play a role in migraine without aura. It is possible that migraine without aura solely depends on vascular changes and neurogenic mechanisms that increase nociceptive receptor sensitization in the brain.
2.1.1.1 Excitatory neurotransmitters and cortical spreading depressionMigraine may be linked to a state of hyperexcitability in brain activity (Ferrari et al., 1990; Petzold et al., 2008). Glutamate and aspartate are the primary excitatory neurotransmitters in the brain, and early studies involving migraine patients showed the levels of glutamate and aspartate were elevated between attacks in patients with migraine with aura as compared to controls (Ferrari et al., 1990). Between these two neurotransmitters, plasma glutamate levels have been found to be elevated over aspartate levels, with each being as high as 0.89 mg/dL and 0.167 mg/dL, respectively (Castillo et al., 1994). Because of this, glutamate has gained the most attention among the two excitatory neurotransmitters and is also seen to be equally high in migraine without aura patients (Zielman et al., 2017). In a preclinical study involving rats, it was observed that there is an increase in glutamate expression which may sensitize neurons in the trigeminal nucleus caudalis (Oshinsky and Luo, 2006). In addition, nociceptive signal transmission in the trigeminovascular system has been shown to involve N-methyl-D-aspartate (NMDA) receptors, and glutamate is an endogenous NMDA receptor agonist (Storer and Goadsby, 1999). Glutamate release and CSD have a bidirectional association in the sense that glutamate stimulates CSD, and this depolarizing wave also enhances glutamate release. In an experiment using rats, CSD was induced by potassium chloride application alone as well as in the presence of the NMDA receptor antagonist MK-801, and MK801-treated brain slices showed an increase in the threshold required for triggering CSD (Petzold et al., 2008). As already mentioned, potassium ion levels increase during CSD activation, but so do glutamate levels and the overall effect is a continuous depolarization of adjacent cells.
2.1.1.2 Ions involved in cortical spreading depressionCortical spreading depression (CSD) has been described to involve an increase in extracellular potassium levels and an increase in intracellular sodium and calcium ions (Leao, 1947; Enger et al., 2015; Cozzolino et al., 2018). Among these ions, change in potassium concentration is the most widely accepted to be implicated in CSD propagation (Tozzi et al., 2012). Grafstein (1956) demonstrated that potassium ions are released by neuronal cells to stimulate a depolarizing cascade in neuronal cells that contributes to the slow propagating wave of CSD (Grafstein, 1956; Somjen, 2001). Additionally, preclinical experiments conducted that involved the measurement of cerebral blood flow and neuronal activity after CSD induction, demonstrated that sodium channel blockers were able to inhibit CSD-induced changes in regional blood flow (Akerman et al., 2008). Topiramate is an antiepileptic with suspected sodium channel blockade activity and used to prevent migraine clinically (Silberstein et al., 2012; Sills and Rogawski, 2020). In rat neocortical slices that have been exposed to increased extracellular potassium ions to trigger CSD, topiramate reduced CSD propagation area as visualized through intrinsic optical signal imaging (Tozzi et al., 2012). The importance of calcium in CSD has also been elucidated, as L-type, N-type and P/Q-type voltage calcium channel blockers reduce repeated CSD onset (Ayata et al., 2000). In addition, mice that express P/Q type calcium channel mutations exhibit higher CSD thresholds induced by electrical and potassium chloride administration (Ayata et al., 2000). Thus, it is evident that calcium channel activation and calcium channel blockers play a critical role in migraine pathophysiology and migraine management.
As discussed above, glutamate is involved in initial CSD generation as well as propagation. Thus, the initiating events surrounding aberrant glutamate release are important in migraine pathophysiology. In a study conducted on migraine patients, magnesium levels in the brain were found to be low (Ramadan et al., 1989) and these findings have led to an interest in magnesium supplementation as part of migraine therapy. Magnesium can inhibit calcium influx, leading to decreased glutamate release and may have other roles such as inhibiting serotonin and calcitonin gene related peptide release, both of which are also contributors to migraine headache (Mathew and Panonnummal, 2021). Thus, it is suspected that low magnesium levels in the brain predispose neurons to aberrantly release glutamate. In support of this hypothesis, an experiment conducted in rats found that local but not systemic magnesium administration inhibited glutamate release as well as suppressed nociceptive responses from the trigeminal region of the brain (Hoffmann et al., 2019).
Migraine headache has long been thought to be due to vasodilation that occurs in meningeal arteries and the next section will discuss the vascular theory of migraine.
2.1.2 Molecules involved in the vascular theory of migraine 2.1.2.1 Nitric oxideNitric oxide is a potent vasodilator that has a causative role in migraine and was part of the molecules investigated to explain the vascular theory of migraine. It was demonstrated in human subjects that an infusion of nitroglycerin, a nitric oxide precursor, was associated with headache of pulsatile quality, and this administration had a ceiling effect at an infusion rate of 0.5 ug/kg/min (Iversen et al., 1989). In another clinical trial on humans, nitric oxide was shown to produce unilateral headache characteristic of migraine with an associated increase in cerebral blood flow due to vasodilation. Quantitative analysis in this study also revealed that, nitric oxide and its metabolites are released in the early phase of a migraine attack (Sarchielli et al., 2000). Not only does its external infusion trigger migraine-like headache, but it was observed that in some cases nitric oxide had the potential to trigger prodromal/premonitory symptoms of migraine (Afridi et al., 2004; Karsan et al., 2016). The action of nitric oxide is one of the major contributors to the maintenance of the vascular theory of migraine because other neuroinflammatory substances discovered that sought to challenge this theory still maintain some association with the release of nitric oxide in the brain (Juhasz et al., 2003).
2.1.2.2 SerotoninSerotonin, a catecholamine also known as 5-hydroxytryptamine (5-HT), has been linked to migraine pathophysiology and some migraine treatments act on the serotonergic system. In a study conducted by Anthony et al. (1967), they observed an apparent significant fall in the plasma levels of serotonin during a migraine with aura attack (Anthony et al., 1967). Similarly, they observed that reserpine administration led to a fall in serotonin levels and triggered a migraine attack (Anthony et al., 1967). Reserpine is a vesicular monoamine transporter-2 inhibitor that blocks the storage of catecholamines and diminishes their concentrations in the synapse (Wimalasena, 2011). As an intervention strategy exogenous serotonin administration reduced migraine headaches. This might also explain why serotonin agonists and medications that increase serotonin levels and activity in the brain such as tricyclic antidepressants, selective serotonin reuptake inhibitors (SSRIs) and serotonin and norepinephrine reuptake inhibitors (SNRIs) have some beneficial role in the treatment of migraine. Studies have also attempted to examine serotonin metabolism during migraine attacks. In one study, contrary to the initial finding by Anthony et al. (1967), serotonin levels during migraine attacks were observed to be higher as well as an observed decrease in 5-hydroxyindoleacetic acid levels (Anthony et al., 1967). This led the authors to conclude leading to a conclusion that serotonin metabolism is elevated during migraine attacks (Ferrari et al., 1989). This conclusion has been corroborated in a study conducted on migraine patients using positron emitted topography scan monitoring 5-HT4 quantification as a surrogate for brain serotonin levels (Deen et al., 2019). In this study, high serotonin levels were detected in correlation with reduced binding to the 5-HT4 receptor. This was determined to be due to 5-HT4 receptor downregulation, and chronic migraine patients were shown to have high levels of serotonin during migraine attack. Thus, serotonin receptors are likely very important in migraine development, maintenance, and treatment.
2.1.2.2.1 Serotonin receptorsThe serotonin receptors 5-HT1B and 5-HT1D have been strongly implicated as having a critical role in migraine pain (Buzzi and Moskowitz, 1991; Tepper et al., 2002). The 5-HT1B receptors mediate vasoconstriction in the cerebral arteries when stimulated by serotonin and its agonists, reversing the vasodilation in arteries to reduce migraine headache. While the 5-HT1B vasoconstrictive effect is also seen in coronary arteries and contributes to its cardiovascular hemodynamic effect, the 5-HT1D receptor is localized within the brain and is involved in inhibiting nociceptive neuropeptide release from the trigeminal system, and minimal vasoconstrictive effects (Goadsby and Hoskin, 1998; Longmore et al., 2000). When stimulated by serotonin 5-HT1B/5-HT1D receptors also inhibit nociceptive signaling between the spinal cord and the brainstem trigeminocervical complex (Tepper et al., 2002). Essentially, these receptors are involved in terminating migraine headache when stimulated and their discovery has led to the drug class known as triptans.
Another class of serotonin receptor, 5-HT1F receptors, are also important in migraine treatment. Unlike the 5-HT1B and 5-HT1D receptors, agonist activity at 5-HT1F receptors does not cause arterial vasoconstriction but instead primarily modulates neurotransmitter release in the brain. The abundance of 5-HT1F receptors in the meninges of the brain, hypothalamus, thalamus and cortex, and presence in trigeminal ganglion neuron terminals allows their ability to block the release of calcitonin gene related peptide (CGRP), as well as glutamate transmission when stimulated by serotonin (Clemow et al., 2020). Because 5-HT1F agonists (ditans) do not produce vasoconstriction, especially in coronary vessels, they do not have hemodynamic side effects and are an attractive therapeutic alternative to 5-HT1B/1D receptor agonists (triptans). The introduction of the peptide calcitonin gene related peptide in this current discussion leads to another theory that deviates from attributing migraine headache solely to vascular mechanisms but also considers how the brain and some associated peptides contribute to migraine headache.
2.1.3 The neurovascular theory of migraine 2.1.3.1 The role of the trigeminovascular system in migrainePrevious studies have shown that certain parts of the brain play a critical role in the pathophysiology of migraine including the trigeminovascular system (Goadsby and Edvinsson, 1994; Stankewitz et al., 2011). The trigeminal nucleus lies in the medulla of the brain stem, and receives sensory input about touch, nociception and temperature from the trigeminal nerve, the facial nerve, the glossopharyngeal nerve, and the vagus nerve. The brain stem has long been associated with migraine triggers, a finding discovered with the aid of functional imaging techniques (Weiller et al., 1995; Bahra et al., 2001). One study that challenged the vascular theory by implicating the potential role of peptides in migraine was conducted by Goadsby et al. (1990) in which an increase in neuropeptide Y, substance P and CGRP in external jugular blood was observed after stimulating the trigeminal ganglion of humans. In another study using cats and humans, trigeminal ganglion stimulation produced CGRP elevations, a response that was attenuated by the anti-migraine drugs sumatriptan and dihydroergotamine (Goadsby and Edvinsson, 1993). The trigeminal nerve which projects into the meninges of the brain was shown to be involved in the migraine attenuation mechanism of serotonin receptors 5-HT1A, 5-HT1B, and 5-HT1D. Agonists of these receptors (triptans) were able to inhibit the electrophysiologic effect of the trigeminal neurons in an isolated superior sagittal sinus (Goadsby and Classey, 2003). Similarly, although nitric oxide, through vasodilatory mechanisms, is able to induce migraine, in humans nitric oxide may activate the trigeminal system through release of peptides and inflammatory substances such as prostaglandin E2 (PGE2), neurokinin A, as well as cAMP (Sarchielli et al., 2000). Magnetic resonance imaging has been conducted on patients who received nitroglycerin infusion and although there was a short-lived vasodilation, unilateral headache persisted with minimal change in cerebral arterial blood flow (Schoonman et al., 2008). The persisting headache can therefore not have been due to vasodilation but through other mechanisms that cause sensitization of nociceptors located in the meningeal regions of the brain. In another experiment using functional magnetic resonance imaging of the brain, a cyclical pattern of activity measured as blood oxygenation level-dependent responses (BOLD), was seen after the trigeminal nuclei were stimulated. A strong BOLD response in the trigeminal nuclei upon stimulation predicted an impending migraine attack and this response was highest in the preictal phase which coincides with the prodromal phase of migraine (Stankewitz et al., 2011). These findings demonstrated an acceptance of neurogenic mechanisms that happen in tandem with vasodilation to explain the pathophysiology of migraine thus strengthening evidence on the neurovascular theory of migraine. But the existing knowledge is not entirely absolute because different markers and receptors come up in migraine research findings that additionally contribute to migraine pathogenesis.
2.1.3.2 Calcitonin gene related peptide—A major molecule associated with the neurovascular theoryAs mentioned above, increased CGRP appears to be correlative with migraine, and is one of such biomarkers that has taken precedence with therapeutic agents being developed against it. CGRP has two isoforms, CGRP-α and CGRP-β (Zaidi et al., 1990; Kee et al., 2018). CGRP was identified to cause vasodilation in cerebral vessels (McCulloch et al., 1986) and subsequently shown to be a part of peptides that are released into circulation during migraine headache (Goadsby et al., 1990; Lassen et al., 2002). This peptide is a 37-amino acid neuropeptide and shown to increase markedly during migraine attacks as demonstrated in a human model of migraine (Juhasz et al., 2003). It acts on a receptor which is a heterodimer of calcitonin receptor-like receptor (CLR) and receptor activity modify protein 1 (RAMP1) to elicit its effects (Liang et al., 2018). It is released by the trigeminovascular system, is expressed by thin unmyelinated C fibers, and it is suspected that activation of these fibers causes CGRP release. However, the receptors CLR and RAMP1 are found in thick myelinated A-fibers (Eftekhari et al., 2013). CGRP has been implicated in preclinical studies using transgenic mice (nestinJ/hRAMP1) as a mediator of photophobia associated with migraine (Recober et al., 2009). The receptor density of RAMP1 has been shown to correlate with the susceptibility of patients to the development of migraine as well (Zhang Z. et al., 2007). The sex-linked pattern of migraine may also be partially attributed to CGRP. In an experiment in which migraine-like response was induced in male and female Sprague-Dawley rats significantly increased hyperalgesic responses were observed in female rats treated with CGRP compared to male rats (Paige et al., 2022).
2.1.3.3 Other biological molecules and structures linked to the neurovascular theory of migraine 2.1.3.3.1 Pituitary adenylate cyclase-activating peptide (PACAP)Pituitary adenylate cyclase-activating peptide (PACAP) is another molecule that is suspected to play a role in migraine. It is structurally related to peptide hormones like glucagon, vasoactive intestinal peptide, and secretin (Harmar et al., 2012). It was isolated in a preclinical rat model and exhibited the ability to stimulate adenylate cyclase release from the anterior pituitary cells (Miyata et al., 1989). PACAP is found in two isoforms known as PACAP-38, the 38-amino acid peptide (Miyata et al., 1989) and PACAP-27, a 27-amino acid peptide, both isolated from rat hypothalamus (Miyata et al., 1990). These two peptides have been shown to induce migraine (Syed et al., 2012; Guo et al., 2016; Ghanizada et al., 2020). Three G-protein coupled receptors are involved in the agonist activity of these PACAP isoforms and they are PACAP type 1 (PAC1) receptor, VPAC1 and VPAC2 receptors (Harmar et al., 2012; Syed et al., 2012). Both PACAP isoforms bind to the PACAP type 1 (PAC1) receptor to stimulate the activation of the trigeminovascular neurons, a phenomenon that leads to vasodilation of meningeal arteries (Akerman and Goadsby, 2015). It must be noted that the role of PACAP in migraine is of emerging importance, but it has been employed in the development of human models of migraine where infusion of PACAP38 triggers migraine-like headache (Schytz et al., 2009). Beyond vasodilation, it is suspected that PACAP i.e., PACAP38 may also cause mast cell degranulation in the dura mater (Baun et al., 2012). It is hypothesized that although vasoactive intestinal peptide (VIP) shares the receptors VPAC1 and VPAC2 with PACAP, it is unable to induce robust degranulation, which might explain why it does not cause headache whereas PACAP activity at VPAC1 and VPAC2 does cause headache (Baun et al., 2012). In addition, sumatriptan, an approved treatment for acute migraine, relieved PACAP38-induced temporal artery dilation and resultant headache (Wienholtz et al., 2021).
2.1.3.3.2 Glial cellsGlial cells are cells found in abundance in the central and peripheral nervous system. In the central nervous system (CNS), the following subtypes are present: astrocytes, oligodendrocytes, NG2-glia and microglia (Ffrench-Constant and Raff, 1986; Kimelberg, 2010; Kimelberg and Nedergaard, 2010; Jäkel and Dimou, 2017; Vila-Pueyo et al., 2023). In the peripheral nervous system (PNS), Schwann cells, satellite glial cells, olfactory ensheathing cells and enteric glia are present (Duncan et al., 1981; Barnett and Riddell, 2004; Rühl et al., 2004; Griffin and Thompson, 2008; Jäkel and Dimou, 2017). Studies have elucidated the role of astrocytes and microglia in the CNS, and satellite cells in the chronification of migraine, CSD initiation and propagation, as well as the symptomology of headache in migraine. Astrocytes are thought to reduce glutamate and potassium ion concentration in the brain using a Na+/K+ ATPase (NKA) pump as well as the glutamate transporters excitatory amino acid transporter 1 and 2 (EAAT1, also known as GLAST, and EAAT2 also known as GLT), respectively (Duan et al., 1999; Capuani et al., 2016; Lee et al., 2022). Mice with a heterozygous mutation that leads to a partial functional loss of the α2 subunit of NKA pump had reduced potassium and glutamate clearance rates (Capuani et al., 2016). In the same study electrophysiological recordings revealed that the brains of these mutant mice also were more susceptible to CSD than wild type mice (Capuani et al., 2016). In a nitroglycerin-induced chronic migraine model using C57BL/6J mice microglia are activated in a proinflammatory manner, and increased proinflammatory cytokine protein is observed (Colonna and Butovsky, 2017; Yang et al., 2023). When roxadustat is administered a reduction in microglial activation and proinflammatory cytokine protein production is observed in the trigeminal nucleus caudalis, as well as attenuated migraine-like behavior (Yang et al., 2023). Satellite glial cells support neurons via cross-communication in peripheral ganglia and are found localized in the trigeminal ganglion (Messlinger et al., 2020). There is also some scientific evidence that shows the satellite glial cells also produce nitric oxide that may contribute to orofacial pain (Vause and Durham, 2009). Neuronal processes are therefore not the only contributing factors to migraine pathology, but there is an apparent augmentation provided by glial cells which requires further exploration.
2.1.3.3.3 HistamineHistamine release has also been identified as a migraine headache contributor. A study conducted by Krabbe and Olesen (1980) showed that infusion of histamine to non-headache, migraine and tension-type headache study participants was able to evoke headache responses in all three groups, and was abolished by the administration of an H1 receptor blocker mepyramine (Krabbe and Olesen, 1980). In a later study, it was shown that the headache response elicited by histamine, due to cerebral artery endothelium activity, was comparable to that seen after nitric oxide administration (Lassen et al., 1995). In addition, mast cell degranulation, also associated with the neurogenic inflammation theory of migraine, causes histamine release (Levy et al., 2007). Histamine release can be also stimulated via CGRP (Ottosson and Edvinsson, 1997). It is hypothesized that brain meninges contain nociceptors which are able to interact with trigeminal neurons, and this trigeminal neuron interaction leads to nociceptor sensitization, ultimately leading to pain, and mast cells contribute to this signaling cascade by releasing serotonin, prostaglandin I2 (PGI2) and histamine (Zhang X. C. et al., 2007). As H1 and H2 receptor antagonism has not been found to produce consistent reductions in migraine headache no antihistamine migraine treatments are currently available (Worm et al., 2019).
Graphical representations of the proposed pathophysiology of migraine are shown in Figures 1, 2. In addition, the prevailing theories of migraine are contrasted in Table 2 while a summary of the mediators of migraine are provided in Table 3.
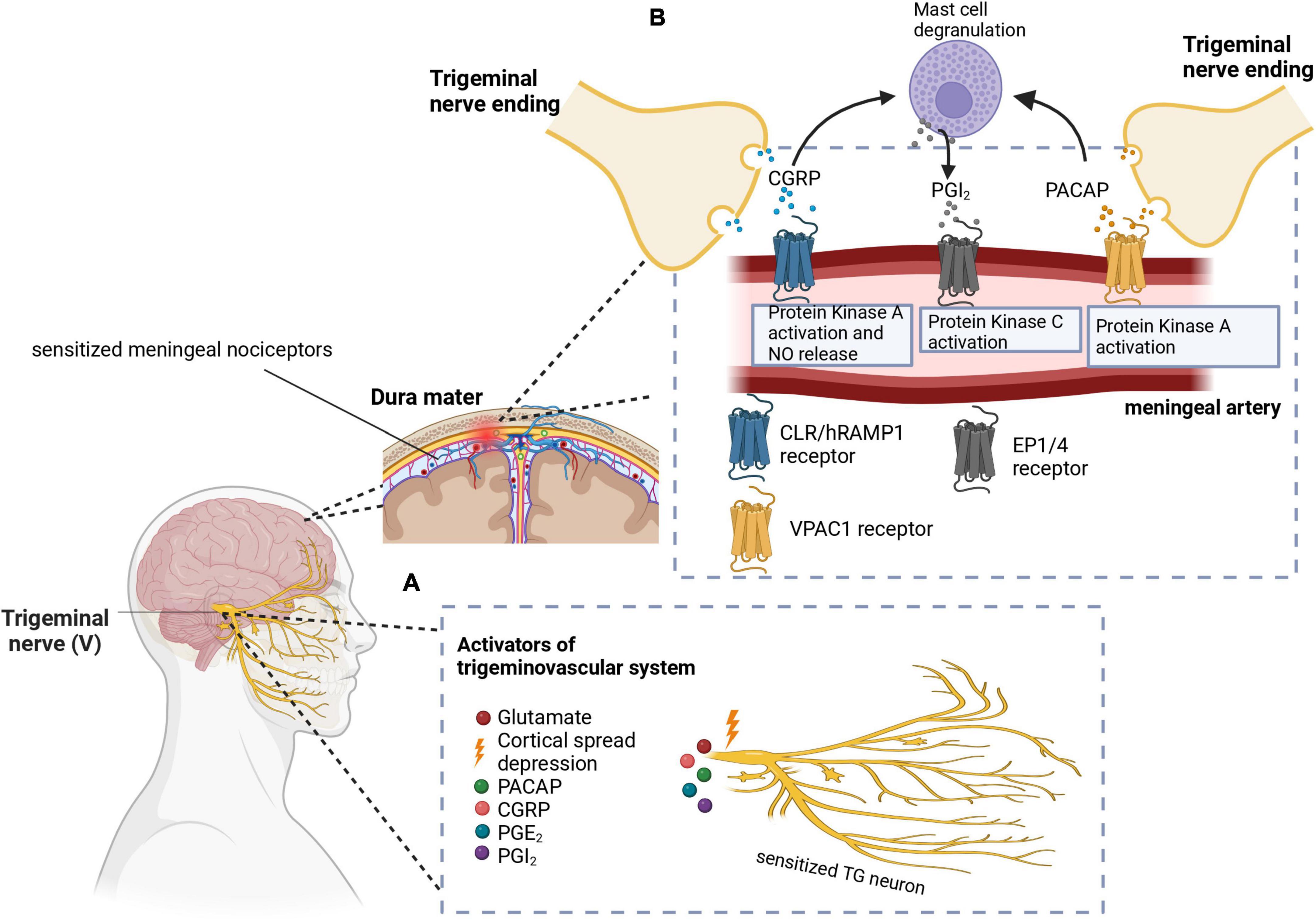
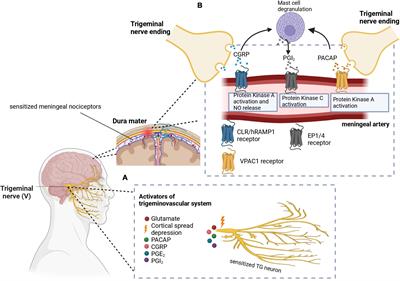
Figure 1. (A) Cortical spreading depression (CSD) stimulates the trigeminal nerve. Additionally, other molecules like PACAP, CGRP, PGE2, and PGI2 have been implicated in the activation and sensitization of the trigeminal nerves. (B) CGRP and PACAP are released from the nerve endings of the trigeminal nerve to cause vasodilation in the meningeal arteries. Signal transduction mechanisms lead to protein kinase A or C activation, as well as nitric oxide release. Mast cell degranulation also produces inflammatory substances such as PGI2 which leads to sensitization of meningeal nociceptors as well as vasodilation. In these images, PACAP, pituitary adenylate cyclase-activating peptide; CGRP, calcitonin gene-related peptide; PGE2, prostaglandin E2; PGI2, prostaglandin I2; CLR/hRAMP1, calcitonin receptor-like receptor/human receptor activity modifying protein 1 heterodimer; EP1/4, prostaglandin E2 receptor. Adapted from BioRender (2023).
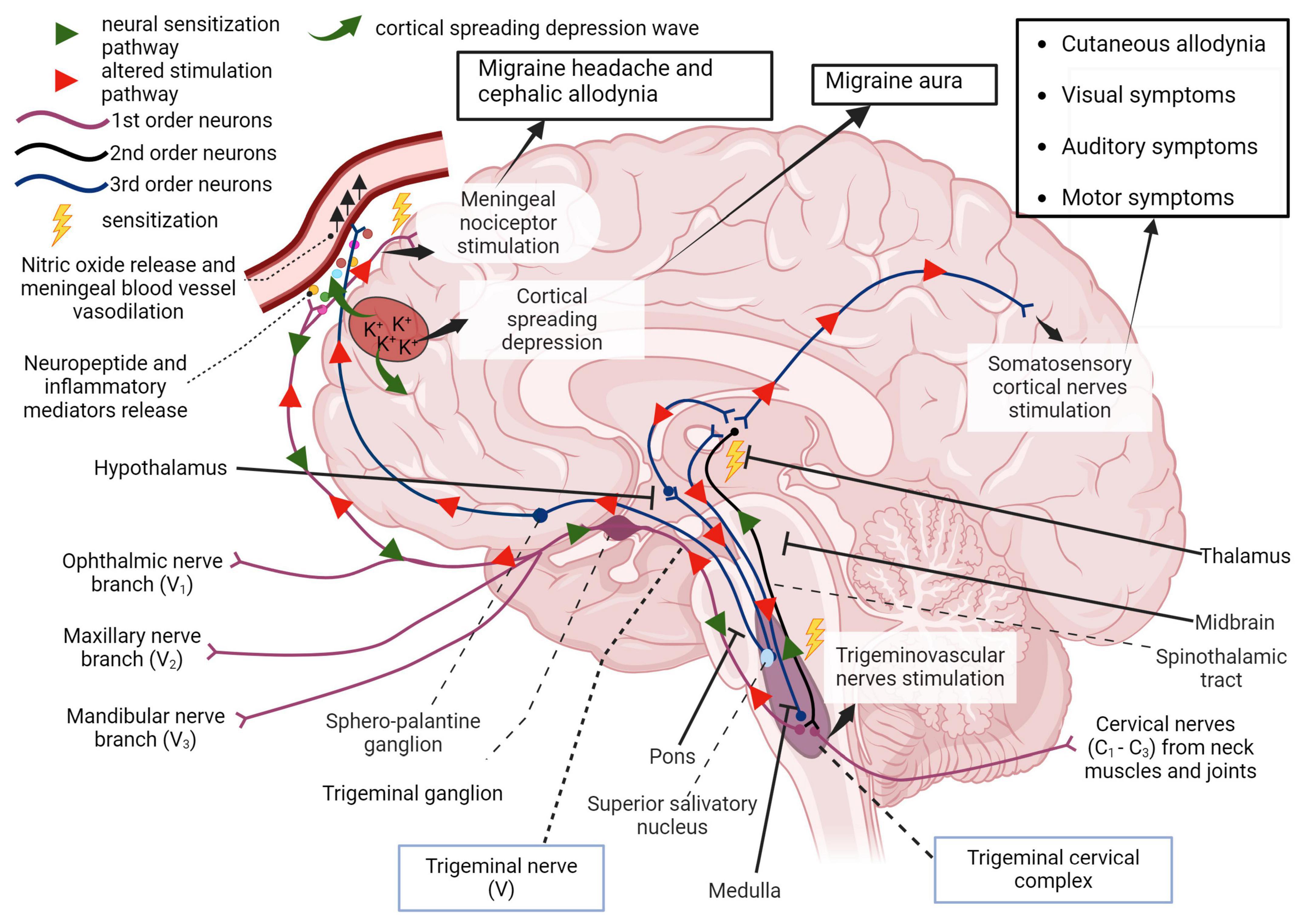
Figure 2. First order afferent nerves of the trigeminal nerve (cranial nerve V) transmit sensory information from the orofacial region and meninges for signal processing in the thalamus. In migraine, peripheral sensitization may occur when neuroinflammatory peptides such as CGRP, as well as processes that lead to mast cell degranulation, cause the downstream sensitization of nerves in the trigeminal pathway. In addition, cortical spreading depression which involves an increase in extracellular potassium ions and enhanced glutamatergic signaling can also trigger this neurogenic response and is also responsible for the symptoms associated with migraine aura. Localized meningeal nociceptor sensitization is another outcome of these processes. The altered afferent signaling pathway sensitizes second order nerves in the trigeminal cervical complex via the trigeminal ganglion and impulses reach the thalamus via the spinothalamic tract. Third order neural projections from the thalamus that innervate the somatosensory cortex, hypothalamus and other areas of the brain undergo an aberrant stimulatory process that causes somatic symptoms of migraine such as extracephalic (cutaneous) allodynia, auditory, visual, and motor impairment. Descending pathways from the thalamus also sensitize the hypothalamus to cause a parasympathetic outflow through the superior salivatory nucleus that promotes nitric oxide release and vasodilation in meningeal blood vessels. The trigeminal nerve endings also secrete neuropeptides that sensitize meningeal nociceptors to cause headache and/or cephalic allodynia (Goadsby et al., 2017; Ashina et al., 2019; Mungoven et al., 2021, 4). Adapted from BioRender (2024).
Table 2. Contrasting the leading theories of migraine pathophysiology.
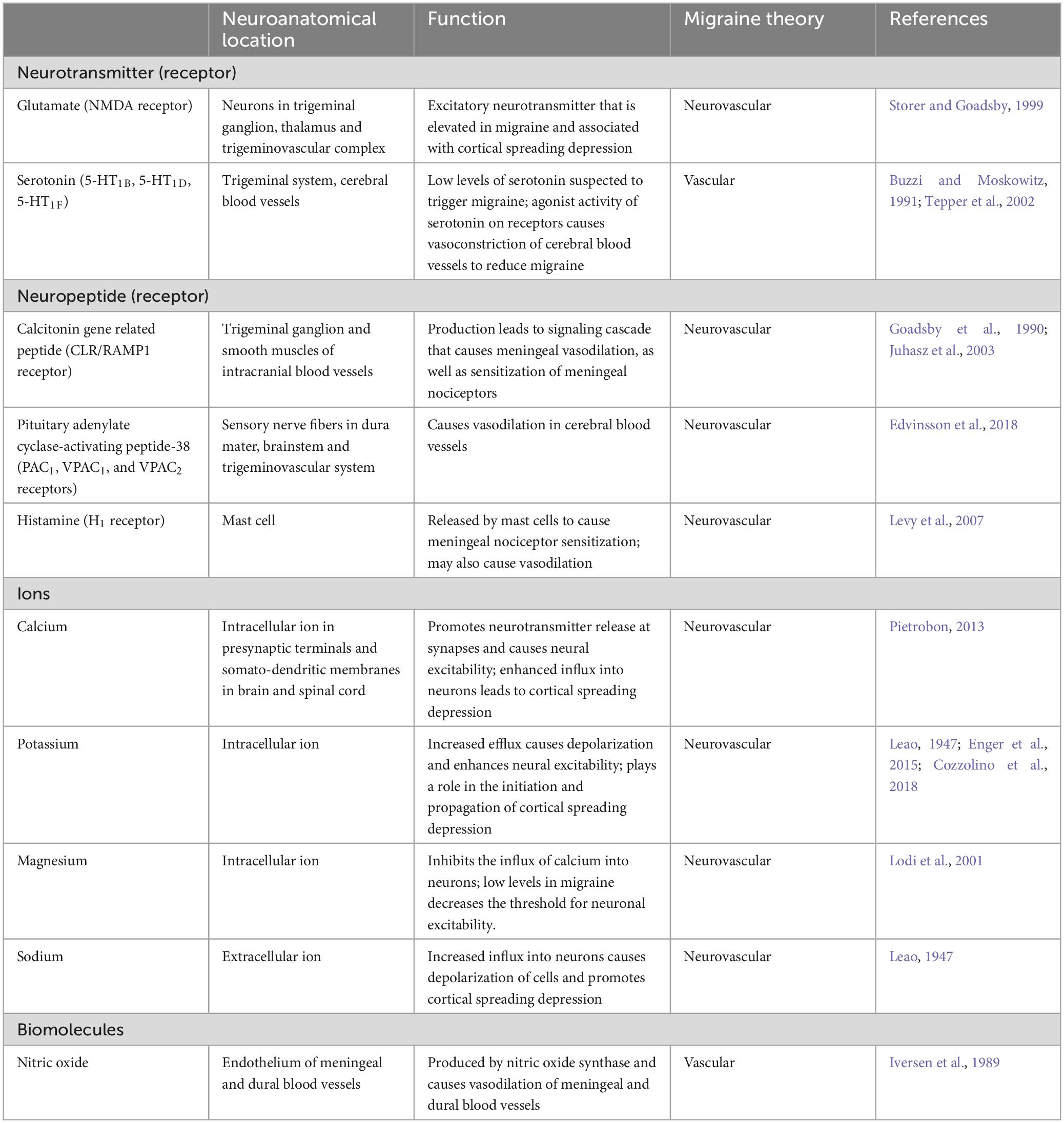
Table 3. Summary of mediators in migraine pathophysiology.
2.1.3.4 Sex-linked hormones in migraine pathophysiologyAbove, it is mentioned that migraine is prevalent in women, and efforts to understand this phenomenon have required investigating the role of menstrual cycle stage and sex hormones in migraine pathophysiology. In a cohort study of premenopausal women with migraine, it was observed that migraine attacks were frequent in the peri menstrual period (2 days before menstruation up to 3 days after the onset of menstruation) with or without aura (Verhagen et al., 2023). A case control study also showed the use of oral contraceptives predisposed patients who experience migraine with aura to increased migraine headaches, and menstruation triggered migraine in patients who suffered from migraine without aura (Granella et al., 2000). Early studies have shown migraine also occurs in pregnancy, especially in patients who experience migraine with aura (Cupini et al., 1995). The estrogen receptor alpha activation, through estrogen agonist activity, is sufficient to produce endothelial nitric oxide (Chen et al., 1999). In preclinical studies, susceptibility to CSD was found to be increased in rats treated with estradiol. Increased CSD due to estradiol may explain the observed increase in migraine with aura among pregnant patients (Chauvel et al., 2018). Immunohistochemistry results also revealed male and female rats do not appear to display differences in CGRP and PACAP, estrogen receptors alpha and beta were more highly expressed alongside CGRP and PACAP in females within the trigeminal ganglion. In addition, in response to estrogen receptor beta stronger middle cerebral artery endothelial vasodilation was observed in female than male rats (Warfvinge et al., 2020).
2.1.4 Genetics in migraine pathophysiologyGenetics plays a role in migraine development as well, and the ICHD-3 document categorizes a rare type of migraine known as familial hemiplegic migraine (FHM) involving migraine aura with motor weakness with a first or second-degree family member having a similar clinical presentation (Headache Classification Committee of the International Headache Society (IHS), 2018). While this form of migraine is not a focus of this review, it is worth mentioning that mutations in the CACNA1A gene, ATP1A2 gene and SCN1A gene lead to the three forms of FHM, respectively, i.e., FHM1, FHM2 and FHM3 (Tottene et al., 2002; De Fusco et al., 2003; Carreño et al., 2013). Migraine with or without aura has been found to have genetic influences as well. One of the early studies conducted to investigate this influence utilized a population-based survey to find out how many migraine patients had relatives who also had migraine. First degree relatives of patients who suffered from migraine with aura had about four times the risk of developing migraine with aura. In a similar manner, first degree relatives of patients who suffered from migraine without aura also had an increased likelihood to develop either migraine with or without aura (Russell and Olesen, 1995). Likewise, a population-based twin study found some level of genetic influence on migraine without aura among monozygotic and dizygotic twins (Gervil et al., 1999). Also, polymorphism in the serotonin transporter transcription gene (5-HTT gene), which is implicated in anxiety related disorders (Lesch et al., 1996), has been associated with inherited migraine. Anxiety is sometimes experienced with migraine and a clinical study showed there was a higher frequency of the s allele of the 5-HTTLPR genotype present in migraine patients (Gonda et al., 2007). The 5-HTTLPR polymorphic variant has also been seen in migraine with aura patients (Borroni et al., 2005). Another finding characterizes the presence of an X-linked dominant allele present on chromosome Xq24-28 that increases the susceptibility to developing migraine (Nyholt et al., 2000). Additionally, ten molecular markers have been detected in the estrogen receptor 1 gene with three of the haplotypes being found to be linked to migraine (Rodriguez-Acevedo et al., 2013).
2.2 United States FDA-approved migraine treatments 2.2.1 Abortive therapiesMigraine treatments fall under two different therapies -abortive or preventive. Abortive medications are taken to stop acute migraines, while preventive medications are taken daily to reduce the occurrence of migraine headaches. A comprehensive review by Zobdeh et al. (2021) classifies all migraine therapies approved by the United States Food and Drug Administration (US FDA) from 1970 to 2020 and a summary is given below. Additionally, in this review newer medications that underwent examination in clinical trials from 2020 to 2023 will be highlighted.
Ergotamine is one of the oldest treatments approved for migraine, but due to its potent vasoconstrictive action its use is hampered by rebound headache as well as a cardiovascular condition contraindication (Tfelt-Hansen et al., 2000). Triptans, serotonin receptor agonists, are the most used drug class used to manage migraines. Some common examples of triptans approved by the US FDA are sumatriptan, zolmitriptan, eletriptan, frovatriptan, naratriptan, rizatriptan, and almotriptan. Zolmitriptan, naratriptan, rizatriptan, almotriptan, frovatriptan are non-selective 5-HT1B and 5-HT1D receptor agonists (Steiner et al., 2003; Marmura et al., 2015; Zobdeh et al., 2021). Eletriptan is a non-selective 5-HT1B, 5-HT1D, and 5-HT1F receptor agonist. Despite their common use in clinical practice, they often produce medication overuse headache, and the discontinuation rate of triptans is between 50 and 82% (Yang et al., 2021), One major contraindication to triptan use is 5-HT1B mediated vasoconstriction which can negatively impact existing cardiovascular conditions. Ditans, 5-HT1F receptor agonists, do not possess the vasoconstrictive activity of triptans, and Lasmiditan is the only member of this class that is FDA approved (Kuca et al., 2018). CGRP receptor antagonists known as gepants are an alternative to triptans and have also been approved to manage acute migraine (Croop et al., 2019). These molecules directly bind to the CLR/RAMP1 receptor to antagonize the pharmacologic effect of CGRP. Rimegepant, ubrogepant, and zavegepant are the current gepants approved by the US FDA for acute migraine treatment (Croop et al., 2019; Dodick et al., 2023; Lipton et al., 2023).
Over-the-counter medications are also used to treat migraine and may be available in combinations. Acetaminophen, caffeine, and non-steroidal anti-inflammatory drugs such as ibuprofen, aspirin, celecoxib are some examples of over-the-counter medicines used for acute migraine (Goldstein et al., 1999, 2006; Zobdeh et al., 2021).
2.2.2 Preventive therapiesFor the prevention of migraine, CGRP human monoclonal antibodies have been developed that either bind CGRP and inhibit CGRP binding to its cognate receptor or directly block CGRP receptors. CGRP-related drugs that have been FDA-approved include erenumab (CGRP receptor blocker), galcanezumab, fremanezumab, and eptinezumab (Silberstein et al., 2017; Detke et al., 2018; Lipton et al., 2020). Other drugs that are FDA-approved for migraine prophylaxis include atogepant (CGRP receptor antagonist), onabotulinum toxin A (acetylcholine release inhibitor), topiramate (sodium channel blocker/glutamate antagonist/GABA modulator), amitriptyline (tricyclic antidepressant), valproic sodium, valproic acid, propranolol (non-selective beta blocker) (Diener et al., 2007; Dodick et al., 2009; Pringsheim et al., 2010; Herd et al., 2018; Steiner et al., 2019; Pozo-Rosich et al., 2023). A summary of approved medications is provided in Table 4.
2.3 Novel and experimental therapeuticsCurrently, LU AG09222 is an investigational monoclonal antibody that targets PACAP38 and has completed a phase 2 clinical trial to evaluate their use in migraine headache prevention (Rasmussen et al., 2023). Cannabidiol in combination with cannabigerol and tetrahydrocannabinol is in a Phase 4 clinical trial to investigate its use as an adjuvant therapy to medication commonly used to prevent migraine (Kaup, 2023). These clinical trials demonstrate that additional targets may prove to be fruitful to examine and may yield novel migraine therapeutics.
In addition to these, some drugs have been repurposed and investigated in clinical trials to be used as monotherapy or combination therapy for acute and chronic migraine. Examples include lidocaine which is a local anesthetic and indomethacin which is an NSAID (Di Monda et al., 2003; Schwenk et al., 2022). Ketamine (NMDA receptor antagonist) and bupivacaine (local anesthetic) are other repurposed drugs also used to manage acute migraine (Afridi et al., 2013; Cady et al., 2015; Cohen et al., 2018). A summary of experimental therapeutics is provided in Table 5.
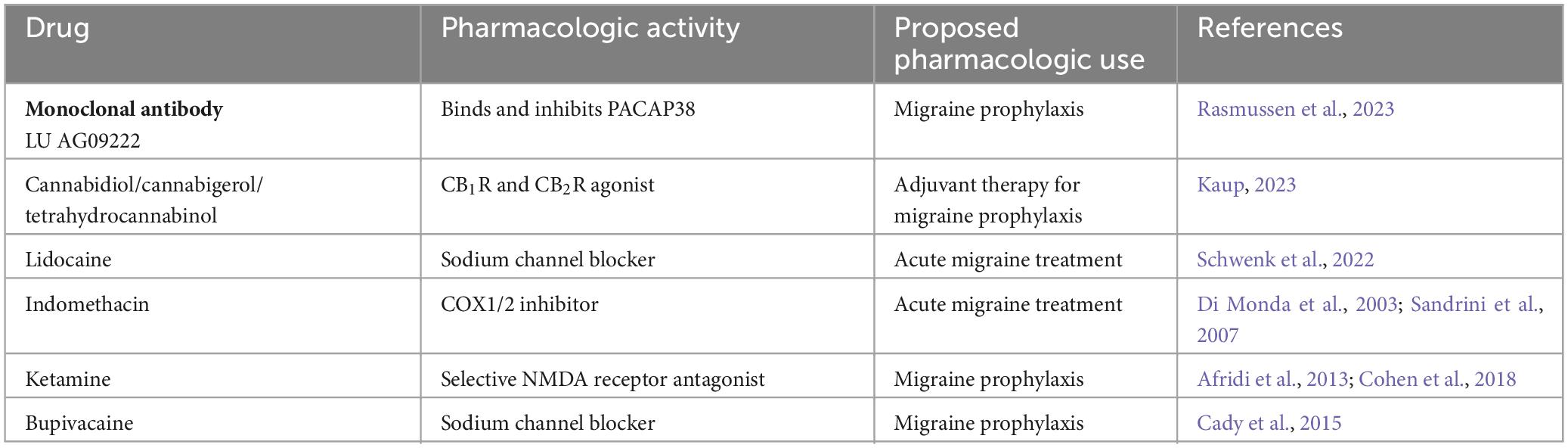
Table 5. Experimental migraine therapeutics.
3 Complications of migraine 3.1 Status migrainosusStatus migrainosus is an intractable and severe form of migraine that persists beyond 72 h and may occur with or without an aura (Headache Classification Committee of the International Headache Society (IHS), 2018). To date, there is no established effective treatment for this migraine complication (Iljazi et al., 2020). This condition, while not fatal, is responsible for frequent hospitalizations and increased healthcare costs. A study by Harnod et al. (2018) also highlighted the tendency for suicide among status migrainosus patients (Harnod et al., 2018).
3.2 Migrainous infarction and strokeMigrainous infarction is a rare migraine complication diagnosed as an ischemic infarction, occurs in a patient experiencing migraine with aura that persists for more than 1 h, and the stroke onset coincides with the migraine attack (Headache Classification Committee of the International Headache Society (IHS), 2018). It must be highlighted that ischemic stroke may occur later in a migraine patient and will not be classified as a migrainous infarct because of its onset. However, stroke is also a migraine complication, with early research suggesting a possible link between the two (Murphy, 1955). Some studies have suggested that migraine could serve as a risk for developing ischemic stroke (Henrich and Horwitz, 1989) but with a higher association among women below 45 years of age (Tzourio et al., 1993).
In addition, magnetic resonance imaging studies point to an increased risk of migraine patients developing subclinical brain infarcts (Monteith et al., 2014) which is associated with higher migraine attack frequency (Kruit et al., 2004). There is evidence that points to migraine patients having ischemic white matter abnormalities (WMAs) seen as hyperintense lesions and suspected to be linked to microvascular damage (Ashina et al., 2021). It has been reported that as much as 59% of migraine patients could have white matter abnormalities (Bashir et al., 2013) and the CAMERA-1 study highlighted a higher prevalence of infratentorial cerebellar infarcts in patients with migraine with aura (Kruit et al., 2005). Compared to women without migraine women with migraine without aura had a higher deep white matter hyperintensity volume (Palm-Meinders et al., 2012). However, no link between WMAs and migraine frequency, migraine severity and migraine type has been found, but these changes may explain why migraine patients are at a risk of stroke-related conditions (Palm-Meinders et al., 2012). There are also studies that highlight an association between migraine and developing cardiovascular disease with relatively higher correlation among patients who experience migraine with aura, especially women (Bigal et al., 2010; Kurth et al., 2020;
留言 (0)