
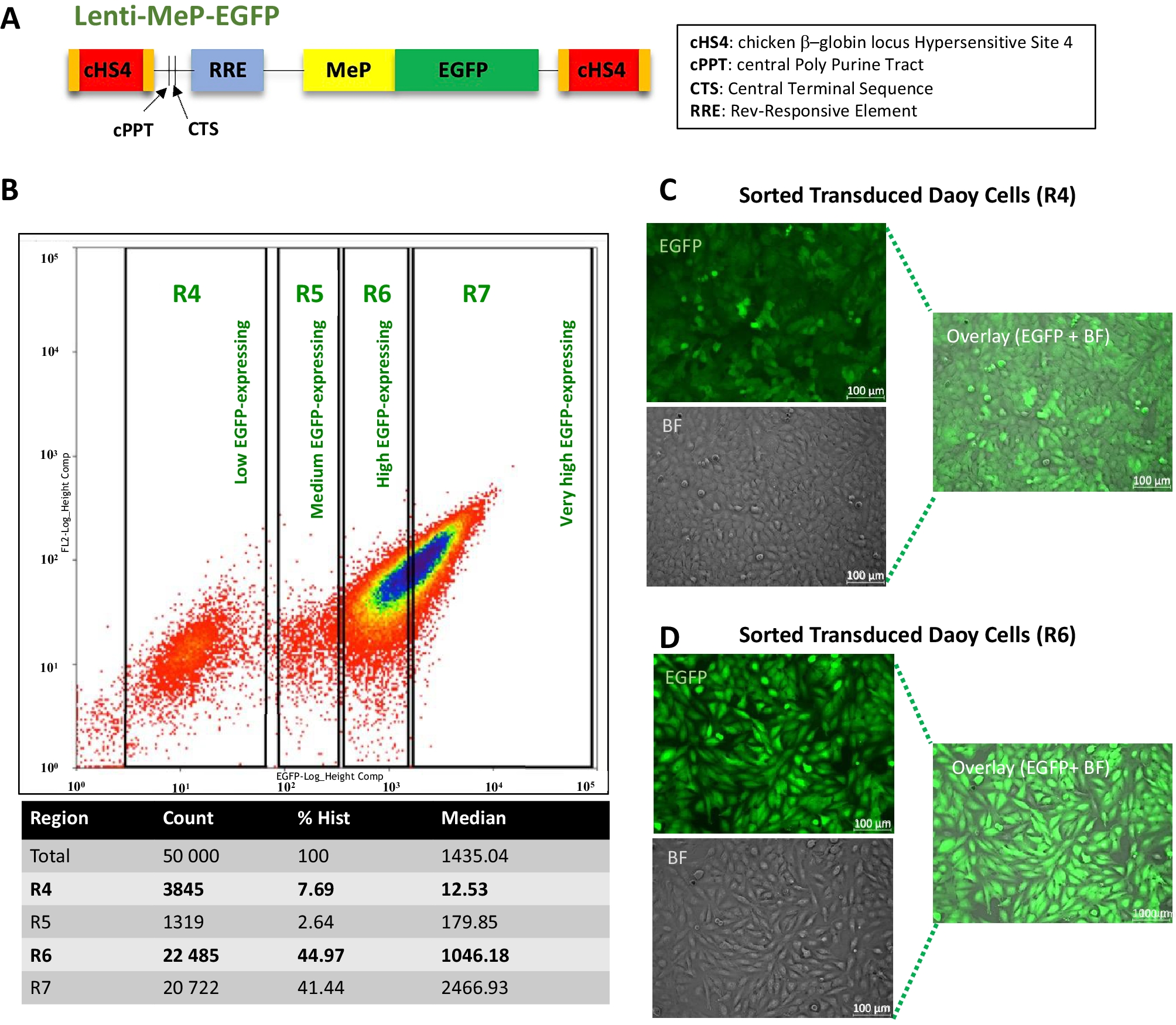
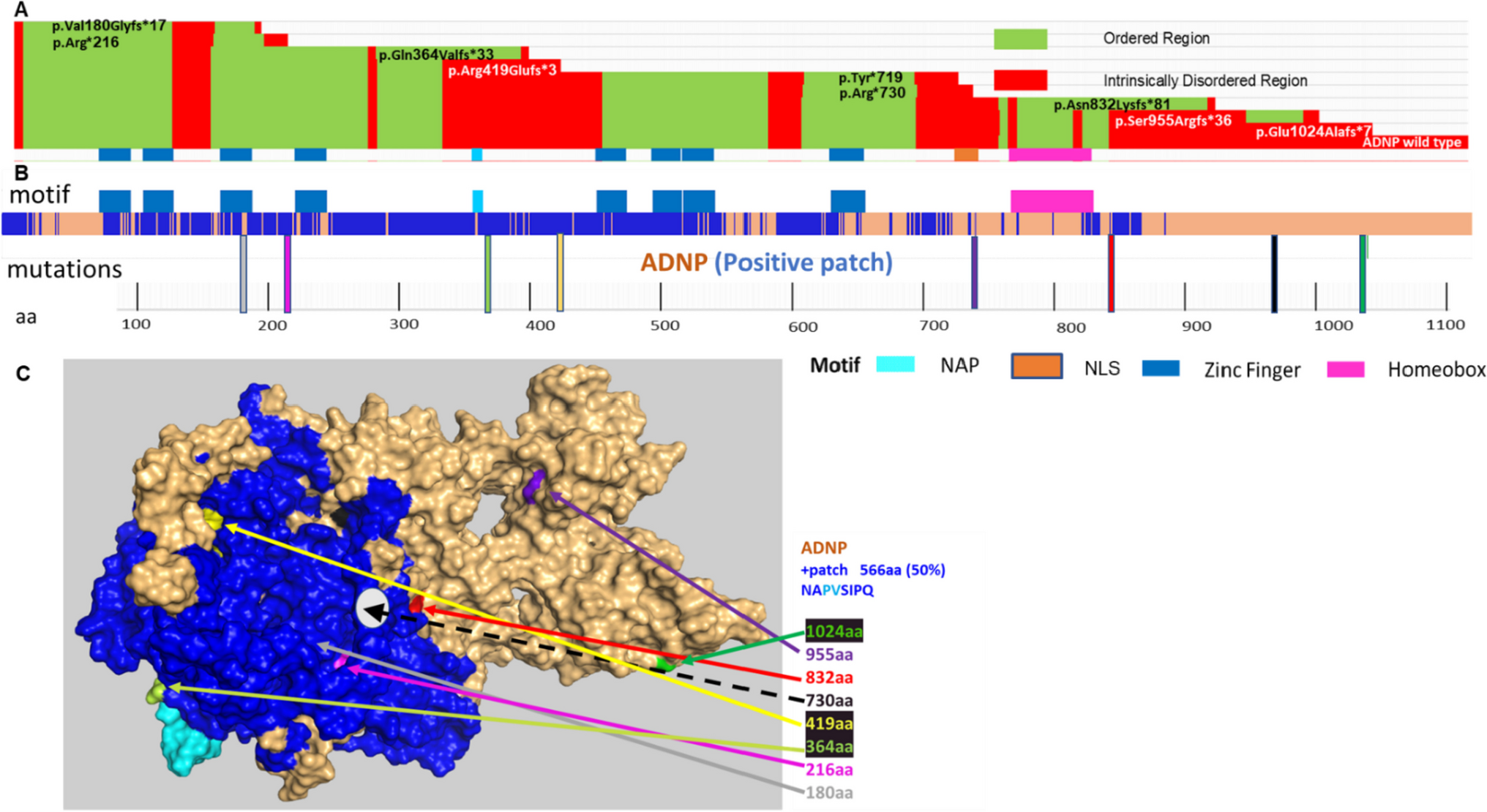
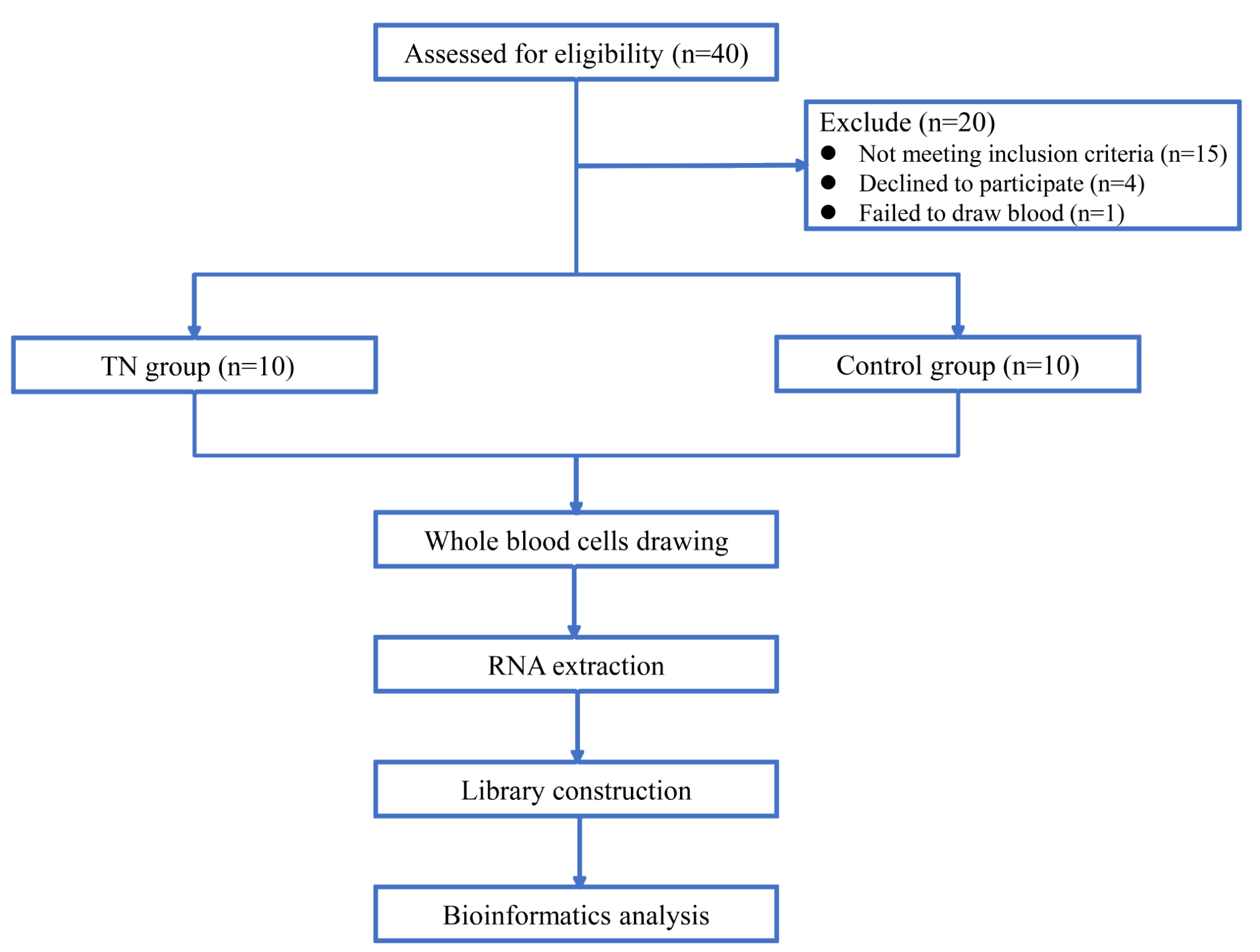
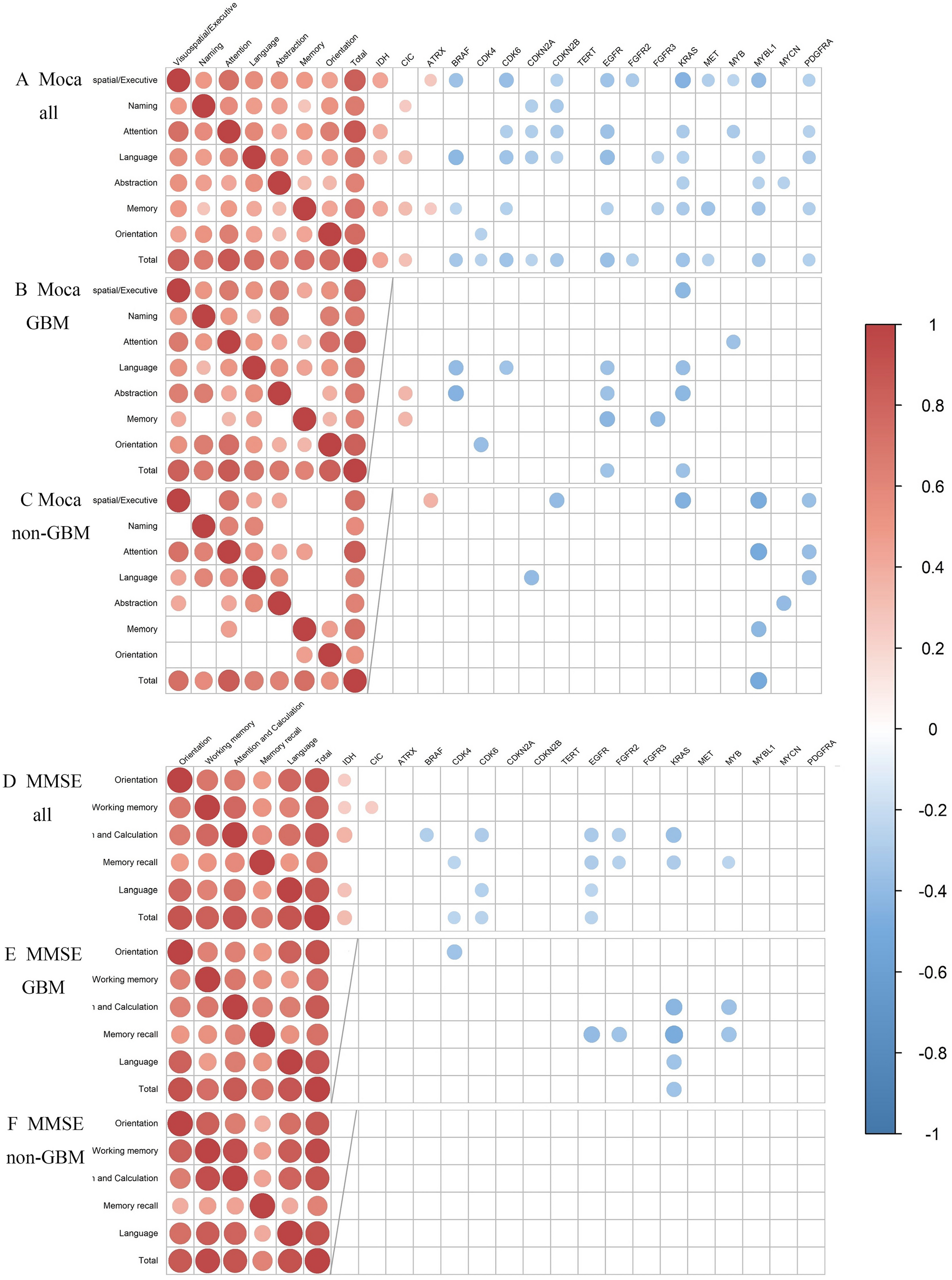
記住我


Ferroptosis is a recently identified type of cell death that differs from apoptosis, necrosis, and autophagy. It has been seen in the neurological system, renal tissue, and cancer cells (Dixon et al. 2012). We focused on hemin-induced ferroptosis in HT22 hippocampal neuronal cells to examine the effects of ferroptosis on brain injury after hemorrhagic stroke in vitro. To investigate the harmful effects of hemin on the HT22 cells, we first used a CCK-8 kit and flow cytometry to measure cell viability and mortality, respectively. After the exposure to different concentrations (5, 25, and 125 µM) of hemin for 6 h, the viability of the HT22 cells decreased (Fig. 2A, P < 0.01, P < 0.0001, respectively), and cell mortality increased with increasing concentrations of hemin (Fig. 2B). The HT22 cells were treated with hemin for 6, 12, and 24 h, and cell viability was approximately 10% at 12 h (Supplementary Fig. 1). In addition, the results showed that cell mortality reached nearly 40% at a concentration of 25 µM after treatment with hemin for 6 h (Fig. 2B), which was used as an appropriate concentration for further studies of ferroptosis mechanisms. At this concentration, ferroptosis occurred without excessive cell death. To observe the viability of HT22 cells more accurately after hemin treatment, we used calcein-AM to label living cells, and the cell viability was observed using laser confocal microscopy. As shown in Fig. 2C, the number of live HT22 cells decreased after hemin treatment, consistent with the quantitative results presented in Fig. 2A and B.
Fig. 2
Toxic effects of hemin on hippocampal neuronal HT22 cells. The HT22 cells were treated with hemin (25 µM) for 6 h. A Cell viability was determined using CCK-8. B Cell mortality determined using PI staining (fluorescence profiles and mean fluorescence intensity (MFI)). C Representative images of the live HT22 cells (Calcein-AM, green) exposed to hemin (25 µM); scale bars represent a magnification of 100× . The values are presented as the mean ± S.E.M., n = 3. *P < 0.05, **P < 0.01, ****P < 0.0001, versus the control group
Hemin Triggers Lipid Peroxide Accumulation and an Adaptive Response to Ferroptosis in HT22 Hippocampal Neuron CellsTo find new methods for reversing hemin-induced toxicity, we concentrated on ferroptosis in neurons, a kind of cell death that is dependent on intracellular iron levels and the formation of lipid peroxides (Gaschler and Stockwell 2017). Additionally, ferroptosis can be prevented by using iron chelators (Dixon et al. 2012). In our study, exposure to hemin (25 µM, for 6 h) significantly increased the lipid ROS and cytosolic ROS content in HT22 cells (Fig. 3A and B). However, pretreating the cells with an iron chelator (DFO) resulted in a significant increase of nearly 25% in the viability of hemin-exposed HT22 cells (Fig. 3F, P < 0.05).
Fig. 3
Effects of hemin on lipid peroxides and ferroptosis-related genes in HT22 hippocampal neuron cells. HT22 cells exposed to hemin (25 µM) for 6 h. A and B lipid ROS and cellular ROS measured using flow cytometry after C11-BODIPY staining and DCFDA staining, respectively. C GSH content was detected using flow cytometry. D and E mRNA expressions of GPX4 and Nrf2 measured using quantitative real-time polymerase chain reaction (PCR). F Cell viability determined using CCK-8. The values are presented as mean ± S.E.M., n = 3. *P < 0.05, ***P < 0.001, ****P < 0.0001, versus the control group; #P < 0.05, versus the hemin-treated group
To further explore the mechanism underlying the frustrated adaptive response to ferroptosis, we probed changes in the mRNA levels of antioxidant enzymes, such as GPX4, glutathione (GSH), and nuclear factor E2 related factor 2 (Nrf2). The GPX4 neutralizes reactive lipids during ferroptosis (Forcina and Dixon 2019), whereas the GSH, a synthetic substrate of GPX4, is required for its lipid neutralization function (Feng and Stockwell 2018). Nrf2, an antioxidant transcription factor, regulates hundreds of antioxidant genes, including GPX4 (Wu et al. 2011). Consistent with the finding of previous research (Alim et al. 2019), the mRNA expression of GPX4 (Fig. 3D, P < 0.0001), GSH (Fig. 3C, P < 0.001), and Nrf2 (Fig. 3E, P < 0.001) was increased in hemin-treated HT22 cells. Collectively, these results suggest that an adaptive response to ferroptosis is triggered after a hemorrhagic stroke in vitro. However, the adaptive response failed to protect HT22 cells from ferroptosis. Based on the established model, we performed transcriptome sequencing for a holistic analysis.
Analysis of Transcriptome Sequencing Database After Hemin ExposureTo thoroughly investigate the mechanism underlying hemin-induced ferroptosis, we analyzed the relationship between the two experimental groups of HT22 cells (with and without hemin) using transcriptome sequencing. Cluster analysis was conducted to examine the similarities between the samples (Fig. 4A). Principal component analysis (PCA) was used to explore the intersample relationships between the two groups (Fig. 4B). In the PCA plot, the expression profiles of the samples exposed to hemin were separated from those of the control group (without treatment).
Fig. 4
Analysis of gene expression profiles after the exposure of HT22 cells to hemin. A Results of cluster analysis. The X-coordinate represents the sample name, the Y-coordinate represents the corresponding sample name, and the color represents the correlation coefficient. B Results of principal component analysis (PCA). C Heatmaps of mRNA expressions in the hemin-treated and control groups. Red represents highly expressed RNAs and blue represents mRNAs with low expression. D The volcano plot and statistic of differentially expressed genes (DEGs) in the control and hemin-treated groups. Gray dots represent RNAs that are not significantly different, and red (upregulated mRNAs) and green (downregulated mRNAs) dots represent RNAs that are significantly different
In addition, differential gene expression analysis (DEG) was performed to identify changes in gene expression between the hemin-treated and control groups (Fig. 4C and D). The gene expression pattern of the hemin-treated group was found to be opposite to that of the control group. As shown in the heatmap (Fig. 4C) and volcano (Fig. 4D) results, a total of 2234 DEGs (∣log2FC∣ > 1 and P < 0.05) were detected in the hemin-treated and control groups, of which 1066 genes were upregulated (upper half of the heatmap) and 1168 genes were downregulated (lower half of the heatmap) after hemin treatment.
Identification of the Target DEGs Associated with FerroptosisTo better understand the genes involved in hemin-induced ferroptosis and their significant differences, the ten differentially expressed mRNAs are presented in Supplementary Table 1 based on the KEGG network of the ferroptosis pathway (Fig. 5). Among them, most genes (Acsl1, Ftl1, Gclc, Gclm, Hmox1, Map1lc3b, Slc7a11, and Slc40a1) were upregulated, whereas Tfrc and Slc39a14 were downregulated. The expression levels of these ten genes are depicted in statistical graphs in Fig. 6A–J.
Fig. 5
KEGG network of ferroptosis pathway based on RNA sequence. The genes in red (Acsl1, Ftl1, Gclc, Gclm, Hmox1, Map1lc3b, Slc7a11, Slc40a1) represent upregulated genes, and the genes in green (Slc39a14, Tfrc) represent downregulated genes after hemin treatment
Fig. 6
Changes in gene levels involved in the ferroptosis pathway after hemin exposure (25 µM, 6 h). A–J Relative mRNA levels of ferroptosis-related key genes associated with KEGG pathway “ferroptosis.” The values are presented as mean ± S.E.M., n = 3. *P < 0.05, **P < 0.01, ****P < 0.0001, versus control group
Analysis of the Functional Enrichment of Ten Targeted DEGs Involved in FerroptosisTo gain a better understanding of the ten targeted DEGs associated with ferroptosis, we performed a GO analysis of those ten genes (Fig. 7). The functional enrichment results revealed 20 significantly enriched terms for biological processes (BP), and negative regulation of cell proliferation, positive regulation of transcription by RNA polymerase II, negative regulation of transcription, and DNA-templated cell death were the most enriched BP terms (Fig. 7A). Additionally, 17 cellular components (CC) items, such as the cytoplasm, autophagosome, and cytoskeleton organization, were significantly enriched, with the cytoplasm being the most enriched CC term (Fig. 7B). Moreover, 19 molecular function (MF) items, including protein binding, metal ion binding, HSP70 protein binding, and regulation of ubiquitin-protein ligase activity, were significantly enriched, with protein binding, metal ion binding, and ATP binding being the most enriched MF terms (Fig. 7C).
Fig. 7
Functional enrichment analysis of DEGs. GO term enrichment analysis of DEGs for the ten target genes involved in the ferroptosis pathway. A BP represents biological process; B CC represents cellular component; C MF represents molecular function
Gene Set Enrichment Analysis (GSEA) Against the Gene Sets of the GO DatabaseTo holistically analyze the effect of hemin on cell function, we performed gene set enrichment analysis (GSEA) based on the gene sets identified in the GO enrichment analysis (Fig. 7). We found that the “glutathione metabolic process” was enriched in the hemin phenotype (Fig. 8A). The glutathione metabolic process was closely associated with ferroptosis (Gao et al. 2015). Notably, Gsto2 (glutathione S-transferase omega 2) is involved in the glutathione metabolic process as an antioxidant enzyme that has a protective role in neuronal cells (Allen et al. 2012). Using the String online tool, we identified ten Gsto2 binding proteins, including GPX4 (Fig. 8B). It has been suggested that Gsto2 has the dual functions of scavenging peroxides and detoxification by catalyzing the binding of GSH to harmful substances (Kim et al. 2017). GSH is the key cosubstrate of GPX4, and the GSH-GPX4 pathway acts as a key regulator in the development of ferroptosis (Friedmann Angeli et al. 2014; Yang et al. 2014). In addition, we detected the expression of Gsto2 by western blotting and found that Gsto2 expression decreased in hemin-treated HT22 cells (Fig. 8C). To confirm that Gsto2 was involved in ferroptotic neuronal damage, we used flow cytometry to detect the lipid ROS level after manipulating Gsto2 expression. As shown in Fig. 8D and E, overexpression of Gsto2 led to a decreased lipid ROS level in hemin-exposed HT22 cells. These results indicated that Gsto2 may play a vital role after intracerebral hemorrhage.
Fig. 8
GSEA against the obtained GO database in hemin-treated HT22 cells and the expression of Gsto2 was detected by western blot. A GSEA plots of the gene sets upon the GO database after hemin treatment. B String PPI analysis of the interaction network of Gsto2. C, D Expression of Gsto2 detected by western blotting. E Level of lipid ROS detected by flow cytometry
Hemorrhage Volume in the Brain Increased and the Expression of Gsto2 Was Decreased After the Hemorrhagic StrokeTo investigate whether Gsto2 is involved in ferroptotic neuronal damage after hemorrhagic stroke, we established a hippocampal hemorrhage mouse model (h-ICH; Fig. 9A and B). As shown in Fig. 9C, Gsto2 expression decreased in the hippocampus 24 h post-injury. This suggests that Gsto2 plays a pivotal role in ferroptotic neuronal injury following hemorrhagic stroke.
Fig. 9
Hemorrhage volume following hemorrhagic stroke and the expression of Gsto2 in the hippocampus. A Representative images of brain sections at 24 h post-injury. B Hemorrhage volume measured in the brain (n = 4). C Expression of Gsto2 in the hippocampus detected by western blotting (n = 4)
留言 (0)