Prenyltransferases are crucial biosynthetic enzymes found in all branches of life. In primary metabolism, prenyltransferases catalyze the head-to-tail coupling of the C5 isoprenoid dimethylallyl diphosphate (DMAPP) and one or more molecules of C5 isopentenyl diphosphate (IPP). Catalysis is initiated by metal-triggered ionization of DMAPP to yield an allyl carbocation that reacts with IPP, and the resulting tertiary carbocation intermediate is quenched by proton elimination to yield C10 geranyl diphosphate (GPP) (Fig. 1) (Davisson et al., 1993, Poulter, 2006). Additional rounds of IPP substitution and proton elimination yield increasingly longer isoprenoids such as C15 farnesyl diphosphate (FPP) and C20 geranylgeranyl diphosphate (GGPP) (Poulter and Rilling, 1978, Kellogg and Poulter, 1997). The length of the final chain-elongation product is determined by the depth of the active site pocket (Tarshis et al., 1996, Chang et al., 2006, Ronnebaum et al., 2021).
In lower organisms such as bacteria, fungi, and plants, as well as simple invertebrate marine animals such as corals, GPP, FPP, and GGPP are utilized as substrates for terpene cyclases to generate aliphatic hydrocarbons, most often with intricate multi-ring structures (Christianson, 2006, Christianson, 2017, Burkhardt et al., 2022, Scesa et al., 2022). In humans, FPP and GGPP are employed in posttranslational modifications to regulate protein interactions with cell membranes, e.g., as found for nuclear lamins A and B, rhodopsin kinase, and the oncogenic GTPase Ras (Zhang & Casey, 1996). Moreover, enzymes involved in protein prenylation can serve as drug targets for the treatment of certain diseases, such as cancer and osteoporosis (Cox et al., 2014, Wang and Casey, 2016, Palsuledesai and Distefano, 2015). Thus, advances in our understanding of prenyltransferase structure–function relationships provide valuable context for current explorations in drug design.
The first prenyltransferase to yield a three-dimensional structure was dimeric avian FPP synthase (Tarshis et al., 1994). FPP synthase adopts an α fold, designated the terpenoid synthase fold (Lesburg et al., 1997, Sacchettini and Poulter, 1997), thought to have evolved from gene duplication and fusion of a primordial 4-helix bundle (Huang et al., 2014). This α fold is shared with class I terpene cyclases (Lesburg et al., 1997, Starks et al., 1997), which similarly initiate catalysis through metal-triggered ionization of an isoprenoid diphosphate substrate. All prenyltransferases contain two signature metal-binding motifs, DDXXD, that coordinate to the catalytically required Mg2+3 cluster (Aaron & Christianson, 2010).
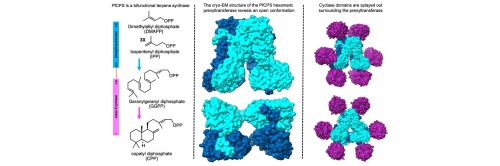
X-ray crystal structures reveal that avian FPP synthase and yeast GGPP synthase form dimers with similar quaternary structures (Tarshis et al., 1994, Chang et al., 2006), but higher order oligomers are observed for prenyltransferases from other species. For example, human GGPP synthase, the GGPP synthase domain of fusicoccadiene synthase from Phomopsis amygdali (PaFS), and the GGPP synthase domain of copalyl diphosphate synthase from Penicillium verruculosum (PvCPS) crystallize as hexamers with D3 symmetry that can be described as trimers of dimers (Kavanagh et al., 2006, Chen et al., 2016, Ronnebaum et al., 2020). The cryo-EM structure of bifunctional macrophomene synthase similarly reveals a hexamer that can be described as a trimer of dimers (Tao et al., 2022). Intriguingly, however, both hexamers and octamers of human GGPP synthase (hGGPPS) are detected in solution (Kuzuguchi et al., 1999), and cryo-EM analysis reveals hexamers and octamers of the GGPP synthase domain of PaFS (Faylo et al., 2021a). Thus, the prenyltransferase dimer can exhibit oligomeric heterogeneity, particularly as hexamers can be in equilibrium with octamers.
Curiously, the crystal structure of D188Y hGGPPS reveals an open hexamer conformation in which one of the three dimer-dimer interfaces is separated, thereby breaking the D3 symmetry observed for the wild-type hexamer (Lisnyansky et al., 2018). The structural basis of this quaternary structural change appears to be a packing interaction with another hexamer in the crystal lattice. It is interesting to speculate that the structure of D188Y hGGPPS represents an intermediate conformation that would be encountered in the hexamer-octamer transition observed for the wild-type enzyme (Kuzuguchi et al., 1999), since the hexamer must open up to allow for insertion of another dimer to form an octamer.
Here, we report the 2.81 Å-resolution cryo-EM structure of the hexameric prenyltransferase (GGPP synthase) core of copalyl diphosphate synthase from Penicillium fellutanum (PfCPS). PfCPS and PvCPS are the first bifunctional terpene synthases to be discovered with prenyltransferase and class II cyclase activities (Mitsuhashi et al., 2017) and are designated as assembly-line terpene synthases (Faylo et al., 2021b, Faylo et al., 2022). In PfCPS, the prenyltransferase α domain is connected to the βγ domains of the class II cyclase by a 69-residue linker (Fig. 2). Surprisingly, the hexameric prenyltransferase core of PfCPS adopts an open conformation such that two dimer pairs are separated, thus breaking the D3 symmetry normally observed for wild-type prenyltransferase hexamers. This open hexamer conformation is similar to that reported for D188Y hGGPPS (Lisnyansky et al., 2018). Even though the class II cyclase domains of PfCPS are randomly splayed out and could not be included in reconstructions of the prenyltransferase core, the structure of the PfCPS prenyltransferase hexamer demonstrates that an open conformation can be achieved even in the absence of a point mutation or interaction with another hexamer.



留言 (0)