Giardia intestinalis is a parasite that causes giardiasis, a diarrheal disease in humans and animals. It is one of the most common pathogens of waterborne transmission; the incidence of the disease is predominant in areas with inadequate sanitary conditions and poor water treatment, where the parasite can be found in its cystic form. The transmission of the disease can also occur by contaminated food. The cysts have a protective wall to survive outside the host. When ingested by the host, the infective cyst differentiates into trophozoites, which colonizes the small intestine. The trophozoite moves around this environment and adheres to intestinal epithelial cells through its microtubular cytoskeletal structures, such as the ventral disc and flagella (Gadelha et al., 2020, Holberton, 1973, Holberton, 1974, House et al., 2011).
Giardia trophozoites have eight flagella, which originate from basal bodies arranged in tetrads, localized between the two oval nuclei in the anterior region of the cell (Elmendorf et al., 2003). Flagellar motility is important for the parasite life cycle because it is involved in the excystation process, cell division, and adhesion to host cells (Buchel et al., 1987, Campanati et al., 2002, de Andrade Rosa et al., 2017, Holberton, 1973, Ishikawa, 2017, Lenaghan Scott et al., 2011). Thus, there is a correlation between the flagellar movements and the establishment and maintenance of the infection. The parasite flagellar structure is conserved compared to flagella of other eukaryotes, presenting nine peripheral pairs (doublets) and a central pair of microtubules containing radial spokes (Sudhakar et al., 2021). The eight flagella are arranged in pairs, differ in position in the cell, and are named anterior, lateral-posterior, ventral, and caudal.
Moreover, a lengthy, non-membrane-bound cytoplasmic region precedes the point where the flagellum exits. The beating pattern of each flagellar pair differs, leading to specific cell movements. Flagella surfaces are similar, except for the ventral flagella, which have membrane projections (Holberton, 1973).
A peculiar feature of G. intestinalis flagella is the presence of elaborated components associated with the inner portion of the axonemes. Following this, the marginal plates, which are part of the ventrolateral flange, and dense rods, are seen connected to axonemes of the anterior flagella (Maia-Brigagão et al., 2013). Other accessory structures can be identified as paraflagellar rods, which extend parallel to the ventral axonemes (Holberton, 1973), and the dense rod associated with the lateral-posterior axonemes (Friend, 1966). The funis, an array of microtubules, extends dorsally and ventrally to the caudal axonemes (Benchimol et al., 2004, Kulda, 1978). Although the composition, organization, and function of these components have not yet been fully understood, they confer distinctive characteristics to each flagellar pair.
Through the examination of isolated flagella, Sundhakar et al. (2021) identified 475 proteins within the flagellar proteome of G. intestinalis. In addition to proteins that integrate the axoneme (tubulins, dynein heavy and light chains, radial spoke proteins), other proteins were identified. Among these are α-giardins, which share homology with human annexins and display calcium-dependent binding to phospholipids (Vahrmann et al., 2008). Pathuri et al., 2009 proposed that α-14 giardin could be associated with the disassembly/assembly of the flagella during parasite differentiation. γ-tubulin and centrin are localized in the basal bodies, indicating that this region would be a microtubule organizing center (MTOCs) (Corrêa et al., 2004, Gadelha et al., 2022, Nohynková et al., 2000). IFT components (IFT-A and IFT-B), BBsome complex, and EB-1 protein are present in the G. intestinalis flagella. Kinesin-2 and 13 are also identified and are involved in the flagellar length control (Dawson et al., 2007, Hoeng et al., 2008). The flagellar proteomes of G. intestinalis exhibited membrane transporters (as ABC, arginin and hexoses tranporters), vesicle transport and edocytosis proteins (as clathrins, adaptins), signaling proteins (as MAP Kinases, Ca2+ transporters) and coiled-coil proteins. Besides that, post-translational modifications such as acetylation, polyglycylation, and glutamylation of tubulin were also detected in the flagella (Boggild et al., 2002, Campanati et al., 1999, Campanati et al., 2003).
Giardia axonemes nucleate from basal bodies within the cell and retain a long cytosolic region before emerging as active flagella. In contrast to other protists, the presence of the transition zone, a specific domain observed between the basal body and the flagellar axoneme and involved in flagellar traffic, was unclear in G. intestinalis. Unlike other protozoa such as Leishmania and Trypanosoma, where the flagellum arises from the base located within the flagellar pocket (Halliday et al., 2021), the region of the emergence of the Giardia flagellum is not accompanied by any membrane specialization. In the externalization region of the G. intestinalis flagellum, only a slight electron-dense material was observed in the plasma membrane (Hoeng et al., 2008).
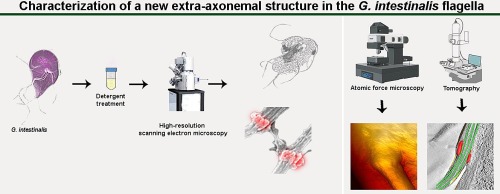
Although molecular analyses of Giardia flagella have been performed using proteomic and biochemical approaches, few studies have focused on the internal organization of the flagella. In this context, our group analyzed the externalization region of the G. intestinalis flagella using ultra-high resolution scanning electron and ion microscopy, atomic force microscopy, freeze fracture, and electron tomography. Our data show that this region has a unique morphological feature: it extends outward, forming a ring-like shape beneath the flagellar membrane. When the plasma membrane is removed, a structure surrounding the axoneme becomes and observed in all flagellar pairs of the trophozoite. This new structure remains attached to the axoneme even when the interconnections between the axonemal microtubules are disrupted due to treatment with a high detergent concentration. Three-dimensional analysis by electron tomography coupled with examination of freeze-fracture replicas provided information on the spatial organization of this structure relative to the flagellar membrane and other flagellar components. Altogether, the results presented herein provide new insights into the structural organization of Giardia flagella and contribute to a better understanding of the cell biology of this parasite. Whether such structure exists in the flagella of other species is currently under investigation in our lab.



留言 (0)