The N-cap and C-cap are amino acid residues that form the first and last hydrogen bonds between the α-helix backbone amide and carbonyl groups (Richardson & Richardson, 1988). In this case, the three residues following the N-cap or preceding the C-cap require the presence of non-helical partners to form such bonds. This is achieved by the formation of capping motifs, many of which have been described.
For the first time, the structures at the N-termini of α-helices were examined in detail by Efimov (Efimov, 1984, Efimov, 1993). The studied βα-arches contain specific hydrophobic interactions between positions i – 1 and i + 4 (βmαn conformation of βα-arches), i – 2 and i + 4 (βmβγβαn conformation), as well as i – 4, i – 1 and i + 4 (βmββαLββαn conformation), if N-cap is i. It was found that when a polypeptide chain enters perpendicularly into an α-helix, the N-cap usually has a β conformation. Bulky hydrophobic residues are forbidden to occupy this position as they cause dehydration of the free amide groups of the backbone.
The Capping box is one of the most famous N-capping motifs. In it, the side chain oxygen of N-cap (occupied by Ser or Thr) forms a hydrogen bond with the backbone amide of i + 3 position (occupied by Glu or Gln), and reciprocally, the side chain oxygen of i + 3 position forms a hydrogen bond with the backbone amide of N-cap (Harper & Rose, 1993).
Other well-known motifs are the Asx-motif and the S/T-motif. They contain two hydrogen bonds and differ only in the N-cap residue: Asp or Asn in the first case and Ser or Thr in the second. The first hydrogen bond is formed between the side chain oxygen of N-cap and the backbone amide of i + 2 or i + 3 position, and the second is formed between the backbone carbonyl group of N-cap and the backbone amide of i + 3 or i + 4 position (Wan and Milner-White, 1999a, Wan and Milner-White, 1999b).
There is also an N-capping motif, in which the N-cap residue is a proline with a sterically strained cis conformation of the amide bond (Dasgupta et al., 2017). The existence of such motifs is especially interesting in the context of this work.
Structures at the C-termini are equally important for stabilizing α-helices. First of all, they occur at α-helices ending in the αL conformation. There are two such motifs: the Schellman motif and the αL motif. In the Schellman motif, the first backbone hydrogen bond is formed between the i + 2 and i – 3 positions, and the second is between the i + 1 and i – 2 positions, if C-cap is i. In the αL motif, such a hydrogen bond is formed between the i + 1 and i – 3 positions. The αL position should be occupied by glycines or residues with flexible side chains (Schellman, 1980, Milner-White, 1988, Efimov, 1993, Aurora et al., 1994).
Also worthy of mention is the Pro C-capping motif, which has a proline residue in the i + 1 position. This increases the occurrence of a number of residues (Asn, Cys, His, Phe, Tyr, Trp, Ile, Val and Leu) in the C-cap position (Prieto & Serrano, 1997).
In addition, specific hydrophobic interactions were found in αβ-arches (Efimov, 1993). They exist between positions i – 4 and i + 3 (αmγαLββn and αmγβαβββn conformation of βα-arches), i – 4, i + 1 and i + 4 (αmγαLβαβββn conformation), as well as i – 4, i + 1 and i + 6 (αmγαLββPααγββn conformation).
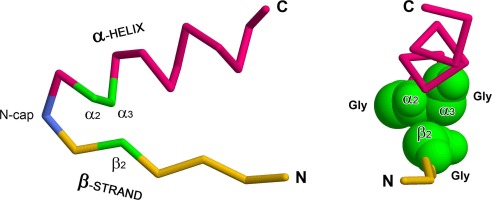
The occurrence of different N-caps may depend on the structural environment. One of the most common supersecondary structures is the βαβ-unit (Fig. 1). It consists of two β-strands, packed in parallel into a β-sheet and connected by one or more α-helices (Rao and Rossmann, 1973, Sternberg and Thornton, 1976). There are right- and left-handed βαβ-units that differ in whether their polypeptide chain is twisted into a right- or left-handed superhelix. Right-handed βαβ-units predominate due to a number of reasons (Rao and Rossmann, 1973, Richardson, 1976, Sternberg and Thornton, 1976, Cole and Bystroff, 2009) based on the homochirality of L-amino acid residues in proteins. Therefore, the left-handed βαβ-units require special conditions for their formation (Kargatov & Efimov, 2020).
Also, a βαβ-unit can be unsplit if its β-strands are adjacent to each other and hydrogen-bonded, or split if one, two or more additional β-strands are packed between them (Orengo and Thornton, 1993, Kargatov, 2020). Each βαβ-unit consists of a βα-arch and an αβ-arch (Efimov, 1993). The first one includes the N-terminal β-strand, N-terminal loop and α-helix, and the second one includes the α-helix, C-terminal loop and C-terminal β-strand. If there are several α-helices, then the ones closest to the N- and C-terminus are considered, respectively. βαβ-units and their constituent βα-arches and αβ-arches are the main structural elements of α/β proteins and are part of many supersecondary structures. Without a complete understanding of their packaging features, it is impossible to model the structure of α/β proteins.
For all amino acid residues, with the exception of glycine, only the regions “α”, “β”, “γ” and “δ” located in the left half of the Ramachandran plot are energetically favorable (Ramachandran et al., 1963, Efimov, 1993). But if in its upper-right quadrant the less favorable region “αL” contains non-glycine residues quite often, then the lower-right quadrant is completely unfavorable for them due to sterical strain.
The present work investigates a novel helical N-capping motif that occurs in βα-arches of βαβ-units. Its N-cap residue has the conformation from the lower-right quadrant of the Ramachandran plot that requires a special structural environment.



留言 (0)