The coronavirus disease 2019 (COVID-19) pandemic caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) has endured for over three years, imposing a substantial social and economic burden worldwide. Despite the development of effective vaccines, a pressing need remains for targeted antiviral therapies against COVID-19 [1]. The invasion of SARS-CoV-2 into host cells encompasses several crucial stages: Initially, the spike glycoprotein of SARS-CoV-2 binds to the ACE2 protein on the cell membrane surface of the host cell. Upon the interaction of the Spike glycoprotein with ACE2, SARS-CoV-2 could enter into the host cell via two distinct mechanisms: 1) Transmembrane protease, serine 2 (TMPRSS2) is responsible for further cleavage and activation the Spike glycoprotein, facilitating the fusion of viral particles with the cell membrane. This fusion allows the release of viral RNA into the cytoplasm, initiating subsequent steps in viral uncoating and replication [2,3]. 2) The ACE2-virus complex can be internalized through endocytosis.Subsequently, mediated by Cathepsin L, viral RNA is released from endosomes into the cytoplasm, instigating infection [4]. In the initial stage of viral infection, SARS-CoV-2 entry into host cells is facilitated by the binding of the receptor-binding domain (RBD) of the spike glycoprotein's S1 subunit to its cell membrane receptor ACE2, triggering endocytosis [5,6].
Coronaviruses employ diverse endocytic mechanisms to access host cells, including CME, caveolae-dependent endocytosis, and clathrin- and caveolae-independent endocytosis. CME is the primary mechanism through which eukaryotes transport cell surface receptors and ligands into cells [7]. Initially, adaptor proteins facilitate clathrin assembly and the cell membrane, forming clathrin-coated pits (CCPs) [8]. Following a maturation process involving various endocytic accessory proteins and dynamic-GTPase action, CCPs loaded with cargos eventually evolve into clathrin-coated vesicles (CCVs) that undergo internalization into the cytoplasm [9,10]. At present, it has been reported that the entry of various coronaviruses, such as porcine epidemic diarrhea virus (PEDV) and infectious bronchitis virus (IBV), depends on the CME process [11,12]. SARS-CoV, the virus mostly similar to SARS-CoV-2, has also been illustrated to utilize CME for cellular entry [13], despite its entrance into the host cell also employing a clathrin-independent process [14]. In the early stages of infection, SARS-CoV-2 has been observed to utilize CME for cellular entry independently of caveolae-mediated endocytosis [4,15]. Consequently, although SARS-CoV-2 might utilize the membrane fusion mechanism mediated by TMPRSS2 to release its genetic material into cells, targeting CME could still be a promising approach to effectively hinder the entry of SARS-CoV-2 and other coronaviruses into host cells, thereby exerting antiviral effects.
In the process of CME, clathrin adaptor proteins form complexes with oligomeric clathrin [16], facilitating its recruitment and assembly into a polyhedral lattice on the plasma membrane. At the same time, adaptor proteins (APs) play a crucial role in mediating the interaction between clathrin and endocytic cargos [17]. Among these, AP-1 and AP-2 are extensively studied adaptor proteins known for their involvement in vesicle formation and cargo transport during endocytosis. Specifically, AP-2 primarily contributes to plasma membrane endocytosis, whereas AP-1 plays a crucial role in the post-Golgi sorting process [18,19]. AP-2, a heterotetramer complex, consists of four subunits: α (AP-2A1), β2 (AP-2B1), μ2 (AP2M1) and σ2 (AP-2S1) [20]. Among these subunits, AP2M1 performs vital functions through the following activities: firstly, AP-2 recognizes and binds the tyrosine based group (YXXØ, Ø indicates a hydrophobic residue) to cargo via AP2M1; secondly, AP-2 possesses three binding sites for phosphatidylinositol-4,5-diphosphate (PIP2), located on AP-2A1, AP-2B1, and AP2M1, respectively, where it interacts with PIP2 in the cell membrane [7]. Recently, an AP2M1 binding site was identified in the cytoplasmic tail of ACE2, which indicated a potential direct interaction between AP-2 and ACE2 [21].
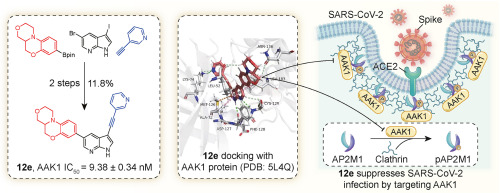
Phosphorylation of threonine 156 (Thr156) on AP2M1 is a critical event in AP-2-mediated CME. When Thr156 of AP2M1 undergoes phosphorylation, it induces a conformational change in AP2M1, facilitating AP-2 binding to the YXXΦ motif, thus internalizing cargos [22]. Two host cell kinases, AAK1 and Cyclin G associated kinase (GAK), catalyze the phosphorylation of AP2M1 on Thr156 [[23], [24], [25]]. Through inducing both the recruitment of clathrin and the phosphorylation on Thr156 residues of AP2M1, AAK1 and GAK enhance the affinity of AP2M1 for binding to cargo proteins on the cell membrane, consequently enhancing cargo endocytosis [[26], [27], [28]]. This characteristic positions AAK1 and GAK as potential molecular targets for the development of antiviral agents. Interfering the function of AAK1 can inhibit the invasion of a variety of viruses into the host, including HCV [29], RABV [30], DENV and EBOV [19]. Recent in silico analysis [31,32] and the latest research by Karim et al. [33] have identified AAK1 as a potential cellular target for SARS-CoV-2 treatment.
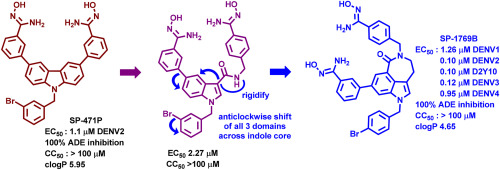
Although AAK1 has emerged as a promising antiviral target, AAK1 has been studied primarily as a drug target for the treatment of neurological disorders [34]. For example, among several known potent AAK1 inhibitors reported in literature (1–9, Fig. 1) [[35], [36], [37], [38], [39], [40], [41], [42], [43]], LX9211 (4) is the clinical stage investigational drug for the treatment of neuropathic pain. Many AAK1 inhibitors have not been assessed and/or optimized for antiviral activity. Previously, pyrrolo[2,3-b]pyridine-based derivative 7 (Fig. 1) was found to have AAK1 inhibitory activity through cross-screening exercise [35]. In 2019, compound 7 and its derivative 8 (Fig. 1) were reported by Jonghe et al. for the potential treatment of DENV and EBOV [42]. Recently, Wang et al. reported that the FDA-approved antitumor drug Sunitinib (9, Fig. 1) could be repurposed for suppressing SARS-CoV-2 infection effectively inhibiting AAK1 kinase [[19], [33], [41], [44], [45]]. Herein, we could like to report the design and synthesis of novel AAK1 inhibitors derived from the promising hit 7. The structure-activity relationship (SAR) as well as the antiviral activity assessment of these new AAK1 inhibitors will be also discussed.

留言 (0)