Translation of genetic information into protein represents one of the most fundamental processes of living organisms. Translation is performed by ribosomes and is divided into four phases: initiation, elongation, termination, and recycling. Despite sharing the same global steps for translation, not all organisms perform translation in the same way, as they have evolved to optimize specific characteristics. Translation initiation is the most divergent and complex process among the different kingdoms of life. It is a highly regulated complex multistep process that requires several initiation factors.1 It plays a key role in ensuring proper positioning and recognition of the start codon by the initiator tRNA: fMet-tRNAfMet and promoting subunit joining to form the 70S elongation-competent complex (70SEC), ready for nascent polypeptide elongation.2 One of the main differences between translation initiation in eukaryotes and prokaryotes lies in the characteristics of their respective mRNAs. Eukaryotes perform a cap-dependent translation initiation, which involves at least nine initiation factors, methionyl-tRNA, and the messenger-RNA (mRNA), which has specific features such as the 5′-cap, the poly-A tail, the Kozak consensus sequence, and the initiation codon which is recognized during scanning of the mRNA.3, 4 Bacterial translation initiation involves three initiation factors and an fMet-tRNAfMet as initiator tRNA, but the mRNA is not scanned. Instead, a ribosome binding site (RBS), located in the 5′ UTR region, is utilized for placing the mRNA into the initiating position. The RBS contains the Shine-Dalgarno (SD) sequence, complementary to the 5′-end of the 16S RNA, or anti-Shine-Dalgarno (aSD) sequence.5, 6 This interaction helps position the initiation codon at the P site, allowing proper recognition of the fMet-tRNAfMet. However, aSD-SD base pairing interaction is not essential for initiation at the correct sites, suggesting that other 5′ mRNA sequences play a role in ribosome alignment as well. In fact, 30–50% of E. coli mRNAs lack the canonical SD sequence.7, 8, 9, 10, 11, 12
An alternative initiation pathway, relevant to this work, has been described for transcripts without a leader, namely leaderless-mRNA (lmRNA), which lacks the upstream 5′ UTR region. These lmRNAs are present in organisms of all kingdoms and are particularly prevalent in Mycobacterium, where they represent up to 26% of its genome.13, 14 Translation of lmRNA can follow canonical initiation or the alternative non-canonical initiation15, 16 in which lmRNA can be directly loaded on the 70S ribosome.17 It should be mentioned that eukaryotic 80S ribosomes have also been shown to perform leaderless initiation in the presence of the initiator tRNA.16, 18
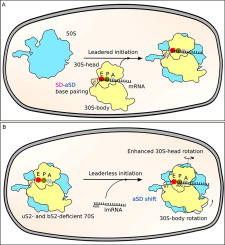
Bacteriophage λ has long served as a model organism for studying gene regulatory mechanisms. The λcI repressor is a critical component of the genetic switch that allows the transition of the phage from lytic to lysogenic development.19, 20 The repressor inhibits λ transcription and thus permits it to maintain lysogeny. In bacteriophage λ lysogens, the λcI repressor is encoded by the leaderless transcript initiated at the λpRM promoter. A down-stream box (DB) sequence (5′-AGCACA-3′) enhances translation of the λcI lmRNA. Host E. coli ribosomes lacking ribosomal protein uS2 translate λcI gene more efficiently than wild-type ribosomes (Figure 1J).21 Without the Shine-Dalgarno sequence, translation initiation can occur either on the mutant 70S or the 30S initiation complexes. E. coli mutants rpsB11 and rpsB1 have improved lmRNA translation efficiency. These were characterized and have been the subject of continuous research.15, 21, 22, 23, 24, 25, 26, 27 So far however, no structural information is available to explain the improved translation efficiency of the λcI lmRNA by uS2-deficient host ribosome. We selected mutant rpsB11 for this study. This mutation is an insertion of the transposable element IS1 next to the uS2 stop codon, which reduces cellular levels of uS2. It only limits the availability of uS2 without modifying the protein and results in two types of ribosomes: those with and those without protein uS2 in the cell. We were therefore able to analyze ribosome structures with and without uS2 in the same sample.
In this work, we investigated the role of ribosomal protein uS2 and the structural features that characterize and explain the improved ability of mutant rpsB11 to translate λcI lmRNA. To this end, we solved the structure of the 70SEC of E. coli mutant rpsB11 loaded with leaderless mRNA and fMet-tRNAfMet (B11), using single-particle cryo-EM. The same complex of 70SEC from E. coli wild-type strain MRE600 (MR600) was used as control. A minimal λcI lmRNA with 12 bases was used, which comprised the λcI DB sequence (5′-AUGAGCACAAAA-3′) shown to be important in the translation of λcI lmRNA.1 This λcI lmRNA spans the P site, the A site, and the entrance channel without protruding from the ribosome. Our cryo-EM structures reveal that the rpsB11 ribosome contains two ribosome populations: (1) 70SEC without uS2 and uS21 (nB11), and (2) MR600-like 70SEC possessing both uS2 and uS21 (wB11). The uS2-deficient ribosomes not only lack bS1, which binds to the ribosome using uS2 as a scaffold, but are also deficient in bS21. Protein bS21 is normally in contact with the aSD, and its absence repositions the aSD, which would otherwise interfere with the positioning of leaderless transcripts in the mRNA exit channel. We find that in nB11 the absence of bS21 and uS2 increases the 30S head dynamics compared to either MR600 or wB11, which produces charge flow and neutralization through the mRNA entrance channel and thereby increases efficiency in translation of λcI lmRNA transcripts.

留言 (0)