As an important source of new drug discovery, natural products play a vital role in drug development [1,2]. According to statistics, approximately 35% of drugs are derived from natural products, with around 25% originating from natural plants [3]. Especially in anti-cancer and anti-infective drugs, the proportion is even higher [3,4]. With increasing interest in developing bioactive ingredients and identifying their molecular mechanisms, several natural derivatives with anticancer effects have continuously emerged in recent decades [4].
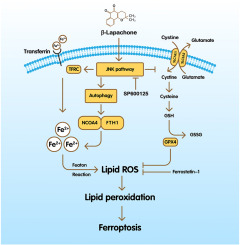
β-Lapachone is a natural ortho-naphthoquinone compound that is extracted from the lapacho tree (Tabebuia avellanedae), which is indigenous to South America [5]. It exhibits multiple biological effects, including anti-inflammatory, anti-fungal, anti-bacterial, and anti-tumor activities [6]. Recently, numerous studies have demonstrated the anti-tumor efficacy of β-Lapachone against various types of cancer, including but not limited to hepatocellular carcinoma, breast cancer, prostate cancer and lung cancer [[7], [8], [9], [10]]. It is worth noting that β-Lapachone injection (named ARQ 501) has completed a number of cancer treatment clinical trials (NCT00102700, NCT00310518, NCT00358930, NCT00524524, NCT00622063, NCT00075933, NCT00099190). The β-Lapachone prodrug (named ARQ 761) has also completed clinical trials (NCT01502800) [11,12], illustrating its great potential in cancer treatment, and in-depth study of its molecular mechanism can guide clinicians to use it more selectively and specifically in clinical practice.
Mechanically, β-Lapachone has a particular mechanism to stimulate NQO1 and shows tumor inhibitory effects by increasing intracellular ROS levels [13,14]. However, the accumulation of ROS has been reported to implicate in lipid peroxidation (LIP) and ferroptosis in cancer cells [15], indicating that β-Lapachone maybe triggers LIP and ferroptosis by increasing intracellular ROS levels. Indeed, the RNA-seq data generated in our study also revealed that the anti-tumor effect of β-Lapachone may be closely related to ferroptosis. Hence, the major forms of death mediated by β-Lapachone remains to be investigated in CRC, especially ferroptosis.
Ferroptosis, differing from necrosis, autophagic cell death, and apoptosis, is caused by the lipid peroxidation of unsaturated fatty acids on the cellular membrane in the presence of intracellular divalent iron or ester oxygenase [16]. Regulation of iron metabolism performs an increasingly important role in ferroptosis. Iron catalyzes the production of ROS and LOXs. LOXs can oxidize cell membrane PUFAs, resulting in the generation of lipid peroxides during ferroptosis [[17], [18], [19]]. Moreover, ferritinophagy exerts crucial effects in the ferroptosis via regulating iron metabolism, especially by inducing ferritin degradation [20]. In the process of ferritinophagy, autophagosomes employ NCOA4 as a selective transport receptor to recognize FTH1, delivering it to the lysosomes for degradation. This process releases free iron ions, leading to an increase in intracellular free iron levels and the accumulation of intracellular ROS, ultimately triggering ferroptosis [21,22]. Regulation of ferritinophagy to control cellular iron levels is considered as a potential therapeutic avenue to suppress cancer progression. To date, there are several FDA-approved drugs that act on different cancers by inducing ferroptosis [23]. Targeting the tumor cell death caused by ferroptosis is a promising new tumor therapy.
JNK pathway, a subgroup of MAP kinases, is primarily activated by stress originating from the tumor microenvironment. JNK pathway has been reported to implicate in multiple biological function, including immune response, cell differentiation, cell proliferation, and programmed cell death, including apoptosis, autophagy and ferroptosis [24]. Studies conducted on HL60 cells utilizing SP600125, a JNK inhibitor, have demonstrated the direct involvement of JNK in ferroptosis [25]. In addition, JNK has been shown to up-regulate the expression several autophagy related genes including ATG5, ATG7, LC3, and Beclin1, in response to specific death promoting stimuli [26]. Numerous studies have demonstrated that the activation of the JNK pathway is closely associated with β-Lapachone-induced tumor cells death [27,28]. However, there are few studies on the regulation of ferritinophagy and iron homeostasis by β-Lapachone and JNK pathway. Therefore, we investigated the involvement of the JNK pathway in β-Lapachone-induced ferroptosis in colorectal cancer within this study.
Here, for the first time, we investigated the involvement of ferroptosis in β-Lapachone-induced cell death in colorectal cancer cells. Further investigation revealed that the mechanism by which β-Lapachone induces ferroptosis and specifically identified that β-Lapachone-induced cell death is associated with iron overload resulting from NCOA4-mediated ferritinophagy. Further, our findings demonstrate the involvement of JNK signaling pathway in β-Lapachone-induced ferroptosis and ferritinophagy in colorectal cancer.







留言 (0)