In the last 30 years, the role of protein topology has emerged with increased interest as new entanglements and loop crossings are discovered. Protein topology refers to the local arrangement of neighboring amino acids and the global three-dimensional structure of the protein. Topology plays a crucial role in understanding the structure-function relationships and can provide insights into the mechanisms of protein folding and molecular recognition integral for the proteomic diversity responsible for sustaining life.
Connolly et al. discovered topological features of entangled protein backbones, where the polypeptide occasionally circles back to form a loop which is then threaded (Connolly et al., 1980). Building on this discovery, the Pierced Lasso Topology (PLT) was discovered in the structure of leptin (Haglund et al., 2012). PLTs are found in proteins containing covalent-loops created by an intramolecular disulfide or isopeptide bond, where the polypeptide backbone is threaded through the macrocycle. The PLT is not unique to leptin and encompasses over 600 proteins represented in all kingdoms of life with 14 conserved biological functions (Simien and Haglund, 2021, Niemyska et al., Nov 2016, Dabrowski-Tumanski et al., 2016). The final entangled topology for disulfide-linked PLTs is formed through three possible mechanisms: (i) Plugging where a free terminus crosses the covalent-loop (Sulkowska et al., 2009), (ii) Slipknotting where an internal residue of the free polypeptide chain crosses the covalent-loop (Sulkowska et al., 2009), and (iii) Belting where the protein folds and the covalent-loop wraps around the threaded element to form the covalent-linker (Simien and Haglund, 2021, Perego and Potestio, 2019). These disulfide-linked PLTs form with the catalysis of an oxidoreductase or at the redox potential in vitro and in vivo.
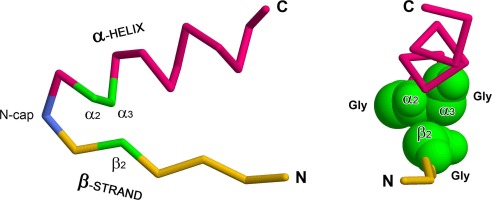
In this work, we utilize the founding member of the PLTs, leptin, to study the folding free energy landscape of the oxidized protein. Leptin is a 146 amino acids protein forming a four-helix bundle motif with a single loop-crossing, a so-called L1 topology (Haglund et al., 2012, Niemyska et al., Nov 2016). Leptin is an ideal protein to study threading as the covalent-loop is 50 amino acids located at the C-terminal. The threaded element is 50 amino acids, leaving 50 amino acids at the N-terminus. Thus, independent of pathway, plugging or slipknotting, there is a 50/50 probability for 50 amino acids to thread through the covalent-loop in either mechanism. However, utilizing the crossing reaction coordinate C (Niemyska et al., Nov 2016) we observe that leptin dominantly slipknots through the covalent-loop 99% of the time (Haglund et al., 2014) (Fig. 1).
In vitro folding experiments show a typical two-state behavior with a linear correlation between the chemical denaturant and the logarithm of the rate for folding (kf) and unfolding (ku) depicted as a chevron plot (Fig. 2). The observed rates are an average of both the plugging and slipknotting mechanisms in the ensemble experiments. In the case of leptin, the observed refolding rate is a direct measurement of the dominant slipknotting mechanism, as plugging only is observed 1% of the time. A previous computational study of a leptin variant, the N-loop protein, has been observed to predominantly plug (Haglund et al., 2017). This loop-variant has two amino acid substitutions increasing the size of the loop to 100 amino acids. The chevron plot shows no significant change in the refolding rate of the N-loop protein where the observed rate is dominated by the plugging mechanism (Fig. 2). Thus, distinguishing the two pathways is complex and requires ingenuity. Molecular dynamics (MD) simulations using the crossing reaction coordinate C are able to support our experimental data to detect loop-crossings. However, the C is not able to detect pre-knotted regions that dictate the threading event.
To further investigate the conformational changes and threading events on the folding trajectories of leptin, the Energy Landscape Visualization Method (ELViM) (Oliveira et al., 2014, Oliveira et al., 2019) is utilized. ELViM is a multidimensional projection tool (Evandro S. Ortigossa et al., 2022, Gareth and Gasparotto, 2019) providing intuitive representations for the energy landscape of biomolecules. Originally designed for visualizing protein folding funnels in lattice models (Oliveira et al., 2014), this method has been successfully extended to more complex biological systems (Oliveira et al., 2019, Sanches et al., 2022, Sanches et al., 2022, Oliveira et al., 2021, Silva et al., 2023). ELViM applies a robust metric to calculate structural distances between pairs of conformations sampled in MD simulations. The dimensionality of the system is reduced to generate an effective two-dimensional representation of the energy landscape that seeks to optimally maintain the original structural distances. The underlying hypothesis of multidimensional projection techniques is that reducing the dimensionality, while retaining relevant information, enables the extraction of significant features that may be concealed in high-dimensional spaces (Evandro S. Ortigossa et al., 2022, Gareth and Gasparotto, 2019, Glielmo et al., 2021). In this study, we utilized ELViM to investigate the folding landscape of wild-type leptin. Analyzing folding trajectories of the wild-type allows for the detection of different pre-threaded conformation associated with the plugging or slipknotting mechanisms within the same structure. The results show that ELViM projection can discriminate the threading mechanisms, where plugging and slipknotting divides the folding landscape into roughly two distinct pathways. In addition, a new reaction coordinate is proposed aiming to classify the propensities of unthread conformations to fold by plugging or slipknotting. As ELViM optimally retains structural similarity in a local neighborhood, the ELViM projection allows the identification of significant pre-knotted regions. Conformations within these regions were utilized to obtain contact maps to shed light on topological clusters that may control the threading mechanism in leptin.



留言 (0)