
記住我
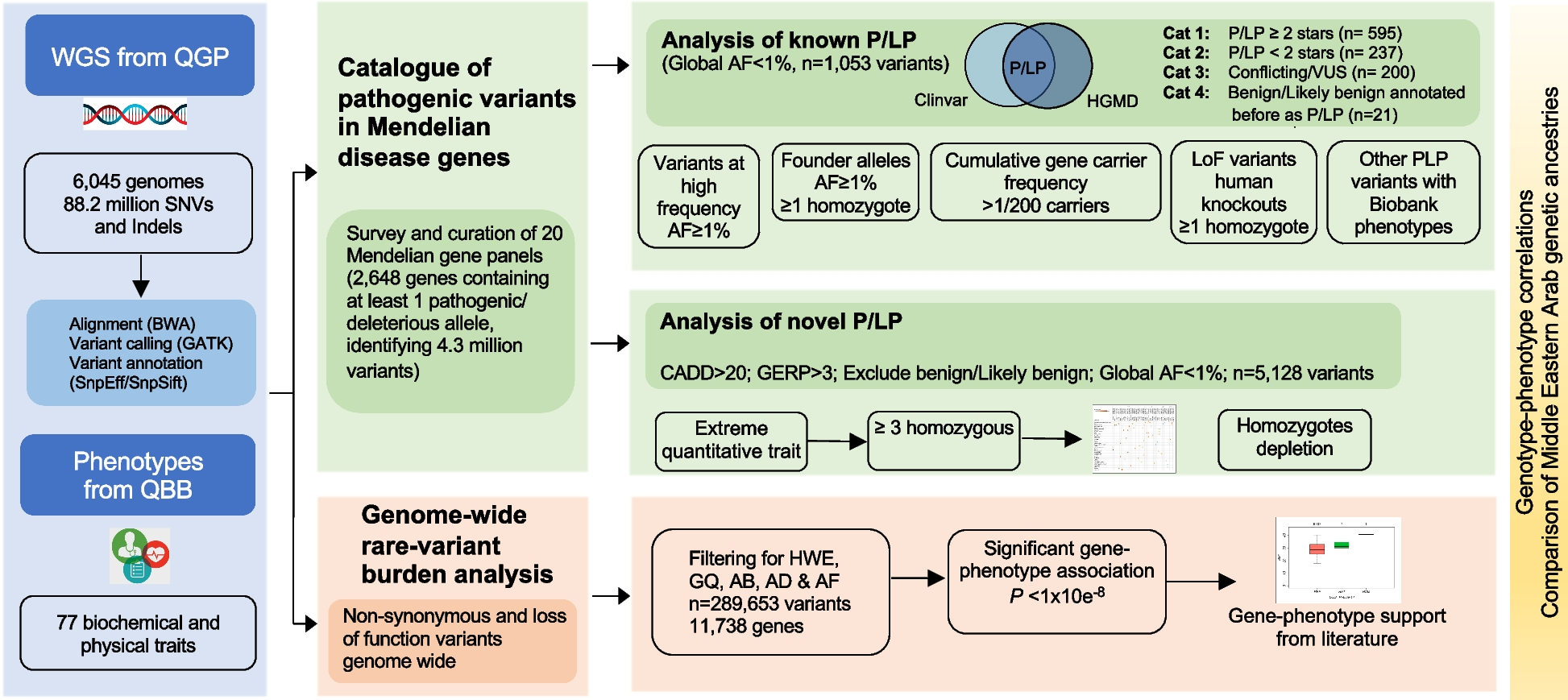





The clinical CRKP isolate HS11286 [44] contains 6 plasmids, including pKPHS1~6 (Fig. 1). The CR IncFII plasmid pKPHS2 (p2) is supposed to be a self-transferable conjugative plasmid, as it carries an oriT, an intact tra gene cluster, a MOBF relaxase gene, and a T4CP gene. The MDR IncA/C plasmid pKPHS3 (p3) is expected to be a nonself-transmissible but mobilizable plasmid, as it lacks the relaxase gene and a complete tra gene cluster but contains an oriT. The other four plasmids, pKPHS1 and pKPHS4~6, are regarded as nonmobilizable. Interestingly, pKPHS3 contains an oriT that shares a similar core nic site with the conjugative plasmid pKPHS2, similar to the mobilizable virulence plasmid pRJF293 (pV) [14] in the K2 hvKP RJF293 (Fig. 2 a). To experimentally investigate the transfer of the conjugative CR plasmid pKPHS2, the mobilizable MDR plasmid pKPHS3 and the mobilizable virulence plasmid pRJF293, we employed CRKP HS11286 to conjugate with hvKP RJF293 and the plasmid-free Klebsiella variicola strain KvBSI002A. In the first round of the conjugation assay, pKPHS2 was transferred from the donor HS11286 to the recipient RJF293 at a frequency of (6.89±1.34)×10−6, and pKPHS3 was transferred at a frequency of (1.29±0.41)×10−8 (Fig. 1; Additional file 6: Table S5). In the second round, pKPHS2 was further transferred into the recipient K. variicola KvBSI002A alone or cotransferred with pKPHS3 and/or pRJF293 (Fig. 1; Additional file 6: Table S5). In this conjugation assay, the transfer of nonmobilizable pKPHS1 and pKPHS4~6 was not detected as expected.
Fig. 1
Transfer of the mobilizable MDR plasmid pKPHS3 (p3) and the virulence plasmid pRJF293 (pV), both lacking relaxase genes, with the help of the conjugative CR plasmid pKPHS2 (p2). a Schematic diagram of the two rounds of conjugation assays. The green square denotes the tra gene cluster on pKPHS2. The green square with white slashes denotes the incomplete tra gene cluster on pKPHS3. The red line indicates the transconjugants generated from K. pneumoniae RJF293-p2 conjugated with K. variicola KvBSI002A. b The conjugation frequencies of pKPHS2, pKPHS3 and pRJF293. The donor strains were HS11286, RJF293-p2, and RJF293-p2-p3. The recipient strains were RJF293 and KvBSI002A. Detailed data are available in Additional file 6: Table S5. c XbaI PFGE and S1-PFGE of transconjugants and their parental strains. M represents the molecular weight marker Salmonella serotype Braenderup H9812. Strains with the same symbol on the PFGE image represent the progeny derived from the same parental strain. The red lines present the transconjugants generated from RJF293-p2 with KvBSI002A. The CR plasmid, MDR plasmid, and virulence plasmid on the S1-PFGE image are indicated by blue triangles, purple triangles, and red triangles, respectively. The plasmids and strains were further confirmed by PCR
Fig. 2
Conjugation assays of gene-edited plasmids. a Alignment of the oriTs of pKPHS2 (p2), pKPHS3 (p3), and pRJF293 (pV). b Schematic diagram of conjugation assays for pKPHS2, pKPHS3, pRJF293 and their nonconjugative derivatives. The green square denotes the tra gene cluster on pKPHS2 (p2). The green square with white slashes denotes the incomplete tra gene cluster on pKPHS3 (pV). The red inverted triangle presents the deletion of the oriT on plasmids. The conjugative frequencies of detected transconjugants that acquired either pKPHS2 (p2) alone or both pKPHS2 (p2) and pRJF293 (p3) are listed. Detailed data are available in Additional file 6: Table S5
To observe the dynamic cotransfer of the mobilizable MDR plasmid pKPHS3 with the conjugative CR plasmid pKPHS2, we selected three K. pneumoniae strains to conduct conjugative assays. The results showed that pKPHS3 can transfer with pKPHS2 from ST11 CRKP HS11286 into K1 hvKP RJF999 and further transfer into K2 hvKP RJF293H (Additional file 5: Fig. S1; Additional file 6: Table S5). The same phenomenon was observed in the mobilizable virulence plasmid pRJF293 (Additional file 5: Fig. S2; Additional file 6: Table S5). When K2 hvKP RJF293-p2 or K1 hvKP RJF999-p2 served as the donor to conjugate with K. variicola KvBSI002A, the mobilizable virulence plasmids were transferred into K. variicola KvBSI002A with the accompaniment of pKPHS2, and further transferred into CRKP HS11286. The cotransfer of mobilizable MDR or virulence plasmids with CR plasmids across different strains generated a series of transconjugants, such as hypervirulent carbapenem-resistant K. pneumoniae (hv-CRKP), and hypervirulent multidrug-resistant carbapenem-resistant K. pneumoniae (hv-MDR-CRKP).
We also examined the cotransmission of the mobilizable MDR plasmid pKPHS3 and virulence plasmid pRJF293 with the conjugative CR plasmid pKPHS2 from K. pneumoniae to E. coli. A typical capsule-free strain, E. coli C600 [45], was employed as the recipient. We found that both mobilizable plasmids pRJF293 and pKPHS3 can transfer with conjugative pKPHS2 from K. pneumoniae RJF293-p2-p3 into E. coli C600. Among E. coli transconjugants, the two mobilizable plasmids stayed separate, or formed hybrid plasmids with pKPHS2 (Additional file 5: Fig. S3; Additional file 6: Table S5). The PFGE and WGS results showed that in transconjugants, pKPHS2 and pKPHS3 formed a 196,720 bp hybrid plasmid p2-3 at the edge of a Tn3 IS family. The fusion event might occur via replicative transposition or homologous recombination (Additional file 5: Fig. S4). In transconjugant C600-p2-V, there were two hybrid plasmids, including a 264,857 bp plasmid p2-V and a 74,014 bp plasmid p2-V-2. These hybrid plasmids speculatively underwent two rounds of recombination events to form a large fusion plasmid derived from pKPHS2 and pRJF293, followed by division into two separate plasmids (Additional file 5: Fig. S5). Since the timing of plasmid fusion events cannot be detected by PFGE or WGS during conjugation, it is unclear whether the fusion occurred before or after plasmid transfer. However, these fusion plasmids remained stable in passage cultures, as confirmed by S1-PFGE (Additional file 5: Fig. S6). We speculate that plasmid cointegration occurred after plasmid transfer and was aimed at adapting the E. coli host, which needs to be verified in the future.
The transfer of mobilizable plasmids depended on their oriT and the aid of the helper conjugative plasmidTo investigate the transmission of the mobilizable MDR pKPHS3 (p3) and the virulence pRJF293 (pV) with their oriTs (Fig. 2 a) and the conjugative CR pKPHS2 (p2), we deleted the relaxase-encoding gene traI from pKPHS2, and then pKPHS2ΔtraI lost its transferable ability. The transfer of pKPHS3 and pRJF293 was also not detected. We also deleted the oriT from pKPHS3, and pKPHS3ΔoriT3 also lost its mobilizable ability (Fig. 2 b; Additional file 6: Table S5). This indicated that the mobility of the MDR plasmid pKPHS3 relied on its oriT and the helper CR plasmid pKPHS2.
We then cloned oriTs of pKPHS2, pKPHS3, and pRJF293 into the vector pACYC184-Apr, respectively. These constructed oriT-containing plasmids rather than the empty vector pACYC184-Apr were transferred from E. coli C600 into K. pneumoniae RJF293H when they coexisted with conjugative pKPHS2. Meanwhile, they failed to transfer when the helper plasmid pKPHS2 was absent. These plasmids were also not transferred under the accompaniment of pKPHS2ΔtraI (Fig. 3; Additional file 6: Table S5). The results showed that the transfer of mobilizable plasmids depended on their oriTs and the aid of the helper conjugative plasmid pKPHS2. Several reports proposed that the transfer of mobilizable plasmids lacking the relaxase gene might require the other plasmids to provide relaxases [3, 46]. Here, we experimentally validated that natural mobilizable MDR or virulence plasmids lacking the relaxase gene in the K. pneumoniae clinical isolates could still be transferred with the aid of the helper conjugative CR plasmid.
Fig. 3
Schematic diagram of the simulation of mobilization of the virulence plasmid pRJF293 (pV) and the MDR plasmid pKPHS3 (p3) with their oriTs and the help of the CR plasmid pKPHS2 (p2). a When accompanied by pKPHS2, pACYC184-oriTp2 can be transferred while the empty vector pACYC184 cannot. b When accompanied by plasmid pKPHS2, pACYC184-oriTp3 and pACYC184-oriTpV can be transferred. c pACYC184-oriTp2, pACYC184-oriTp3, and pACYC184-oriTpV cannot be transferred when accompanied by plasmid pKPHS2ΔtraI. d pACYC184-oriTp3 and pACYC184-oriTpV cannot be transferred alone. The orange circle denotes the backbone of plasmid pACYC184-Apr, and the blue circle denotes the conjugative plasmid pKPHS2. The green square denotes the tra gene cluster. The blue square denotes the oriT derived from plasmid pKPHS2. The purple square denotes the oriT derived from pKPHS3. The red square denotes the oriT derived from pRJF293. The conjugative frequencies of detected transconjugants that acquired either pKPHS2 (p2) or both pKPHS2 (p2) and pACYC184 derivatives are listed. Detailed data are available in Additional file 6: Table S5
Mobilizable plasmids were not affected by the CRISPR–Cas system in recipientsWe found that the conjugative CR plasmid pKPHS2 carries a proto-spacer targeted by the CRISPR–Cas system of the ST15 plasmid-free K. pneumoniae strain KpBSI083A. However, the mobilizable plasmids pKPHS3 and pRJF293 both do not. To study the impact of the CRISPR–Cas system on the transfer of its targeted conjugative plasmid and nontargeted mobilizable plasmids, we used transconjugants RJF293-p2 and RJF293-p2-p3 as donors to conjugate with K. pneumoniae KpBSI083A and its derivative KpBSI083Δcas3H, which lacks the core nuclease Cas3 of the CRISPR–Cas system. The conjugation assays showed that the transfer of pKPHS2 into KpBSI083A occurred at a significantly higher frequency than its transfer into KpBSI083Δcas3H [(1.66±0.09)×10-5 versus (2.44±0.17)×10-6, p = 0.0000097]. Both mobilizable plasmids pKPHS3 and pRJF293 were transferred into KpBSI083A at frequencies comparable with their transfer into KpBSI083Δcas3H [(4.11±3.29)×10−9 versus (2.92±2.38)×10-9, p = 0.90 and (6.78±0.69)×10−9 versus (7.67±3.77)×10−9, p = 0.21], respectively (Fig. 4 a; Additional file 5: Fig. S7; Additional file 6: Table S5). The results showed that the CRISPR–Cas system in the recipient strain could restrain the acquisition of its targeted conjugative plasmid pKPHS2 but not the nontargeted mobilizable plasmids pKPHS3 and pRJF293.
Fig. 4
Effect of the CRISPR–Cas system in recipient strains on the stability of mobilizable plasmids. a The conjugation frequencies of the targeted conjugative pKPHS2 (p2), the nontargeted mobilizable pKPHS3 (p3) and pRJF293 (pV). The donor strain was K. pneumoniae RJF293-p2-p3 and the recipient strains were K. pneumoniae KpBSI083A and KpBSI083Δcas3H. Detailed data are available in Additional file 6: Table S5. b Schematic of plasmid stability in transconjugants during 13 days of serial passage culture. The plasmid was detected by PCR and antimicrobial susceptibility tests. Plasmids that were retained are indicated by gray squares, while plasmids that were lost are indicated by white squares. Full details of plasmid stability in transconjugants are shown in Additional file 7: Table S6. c The proposed mode of the impact of the CRISPR–Cas system on the dissemination of targeted conjugative plasmids
We then examined the genetic stability of these plasmids in the transconjugants of K. pneumoniae KpBSI083A and KpBSI083Δcas3H. We constructed a vector, pACYC184-spacerp2, containing the same proto-spacer as pKPHS2, which was targeted by the CRISPR–Cas system of K. pneumoniae KpBSI083A. After approximately 7 days of serial passage culture without the addition of additional antibiotics, pKPHS2, and the vector pACYC184-spacerp2 were lost in the host strain KpBSI083A. The loss of the plasmid was confirmed by PCR and antibiotic susceptibility tests. Nontargeted pKPHS3, pRJF293, and empty vector pACYC184-Apr all remained stable in KpBSI083A after 13 days of serial passage culture (Fig. 4 b, Additional file 7: Table S6). Meanwhile, regardless of whether the plasmid contained the protospacer, the plasmids all remained stable in KpBSI083Δcas3H in 13-day serial culture (Additional file 7: Table S6). The results showed that the CRISPR–Cas system in the recipient could interfere with the genetic stability of its targeted conjugative plasmid, while the mobilizable plasmids without proto-spacer could be free from interference (Fig. 4 c).
The plasmid distribution in transconjugants also corresponds with the conjugation assay results. When KpBSI083A served as the recipient, we detected two types of new transconjugants, KpBSI083A-p3 and KpBSI083A-pV, which only contained either the mobilizable MDR plasmid pKPHS3 or the virulence plasmid pRJF293. The corresponding transconjugants with only the mobilizable plasmid were not observed when KpBSI083Δcas3H served as the recipient (Additional file 5: Fig. S7). The transconjugants KpBSI083A-p3 and KpBSI083A-pV might result from the degradation of pKPHS2 in KpBSI083A-p2-p3 and KpBSI083A-p2-pV, which is caused by the CRISPR–Cas system (Additional file 5: Fig. S8).
We further compared the proto-spacers of the CRISPR–Cas system in mobilizable plasmids and conjugative plasmids of K. pneumoniae to explore the effect of CRISPR–Cas restriction on the transfer of mobilizable plasmids. We compiled a dataset of 1194 completely sequenced K. pneumoniae genomes containing 3923 plasmids from GenBank (Additional file 6: Table. S5). According to the conjugative mechanism and plasmid mobility, 3,923 plasmids were divided into conjugative plasmids (n = 1073), mobilizable plasmids (n = 811), and nonmobilizable plasmids (n = 2039). In 1194 K. pneumoniae strains, 297 CRISPR–Cas systems were identified and mainly distributed in the sequence types (STs) ST15, ST23, ST147, ST14, and ST45 (Additional file 5: Fig. S9). Most of the CRISPR–Cas systems belong to Types I-E (n = 198), while a few are classified as Types IV (n = 48), V (n = 46), and VI (n = 4), consistent with previous reports [47,48,49,50]. According to sequence similarity, spacers can be divided into 917 unique spacers, including 184 spacers targeting plasmids in K. pneumoniae. Of the proto-spacers located on plasmids, 62.8% (12,922/20,580) and 28.2% (5798/20,580) were found on conjugative plasmids and nonmobilizable plasmids, respectively, while only 9.0% (1860/20,580) were carried by mobilizable plasmids (Additional file 4: Table S4; Additional file 5: Fig. S10). This indicated that the mobilizable plasmids with fewer proto-spacers might have a larger potential capacity to disseminate with a wider host range than conjugative plasmids.
Mobilizable plasmids and conjugative plasmids in K. pneumoniae enriched AMR and virulence genesA number of mobilizable plasmids and conjugative plasmids, including those of considerable size, were distributed in K. pneumoniae of six typical STs from ST11 to ST147 (Additional file 5: Fig. S11). After in silico analysis of the cargo protein-coding genes in 3923 plasmids of 1194 completely sequenced K. pneumoniae genomes, 87.1% (9545/10,954) of AMR genes and 86.8% (347/400) of virulence genes were located on plasmids (Additional file 3: Table S3). In these plasmid-borne genes, 50.1% (4781/9545), 21.4% (2040/9545), and 28.5% (2724/9545) of AMR genes were clustered on conjugative plasmids, mobilizable plasmids, and nonmobilizable plasmids, respectively. In addition, 75.8% (263/347) and 19.9% (69/347) of virulence genes were carried by mobilizable plasmids and conjugative plasmids, respectively (Fig. 5; Additional file 3: Table S3; Additional file 5: Fig. S9). Only 4.3% (15/347) of virulence genes were located on nonmobilizable plasmids. The ST11 clonal strains [17] were the most predominant K. pneumoniae isolates in China and contained these resistance and/or virulence plasmids (Additional file 5: Fig. S12). In ST11 strains, 30.1% (217/720) of plasmids were conjugative and 32.9% (237/720) were mobilizable (Additional file 3: Table S3; Additional file 5: Fig. S9). The above results suggested that in K. pneumoniae, conjugative plasmids, mobilizable plasmids, and nonmobilizable plasmids all carry AMR genes. Meanwhile, mobilizable plasmids are the main carriers of acquired virulence genes in K. pneumoniae.
Fig. 5
Distribution of AMR genes and virulence genes in K. pneumoniae on plasmids with different transmissible abilities. The 1194 completely sequenced K. pneumoniae genomes containing 3923 plasmids were taken from GenBank. The left bars represent the number of AMR genes or virulence genes and their distribution on plasmids with different mobilities. The replicon distribution in plasmids carrying different AMR genes or virulence genes with different mobilities is illustrated via pie charts. Inc. in Replicon presents the sum of replicons ranging from IncI to IncX. Col. in Replicon represents the sum of replicons ranging from ColRNAI to Col156. The remaining replicons represent the unknown replicons detected by PlasmidFinder
As the transfer of mobilizable plasmids requires the aid of conjugative plasmids, we also examined the coexistence of the mobilizable and conjugative plasmids in 1194 completely sequenced K. pneumoniae strains. Of the mobilizable plasmids, 68.7% (557/811) were accompanied by conjugative plasmids (Additional file 5: Fig. S13a). In K. pneumoniae strains belonging to STs associated with the top ten highest number of plasmids, approximately 72.4% of mobilizable plasmids coexisted with conjugative plasmids (Additional file 5: Fig. S13b). The plasmid pattern of prevailing and clinically worrying CRKP strains was further surveyed. Among 537 CRKP strains, 70.5% (423/600) of CR plasmids possessed transmissible ability, and over half of the CRKP strains contained transmissible MDR plasmids. The virulence plasmids in hv-CRKP strains and hv-MDR-CRKP strains were mainly mobilizable and coexisted with conjugative MDR plasmids (Additional file 5: Fig. S13c; Additional file 3: Table S3). This indicated that most of the mobilizable plasmids might have transfer potential with the help of coexisting conjugative plasmids.
留言 (0)