
Remember me
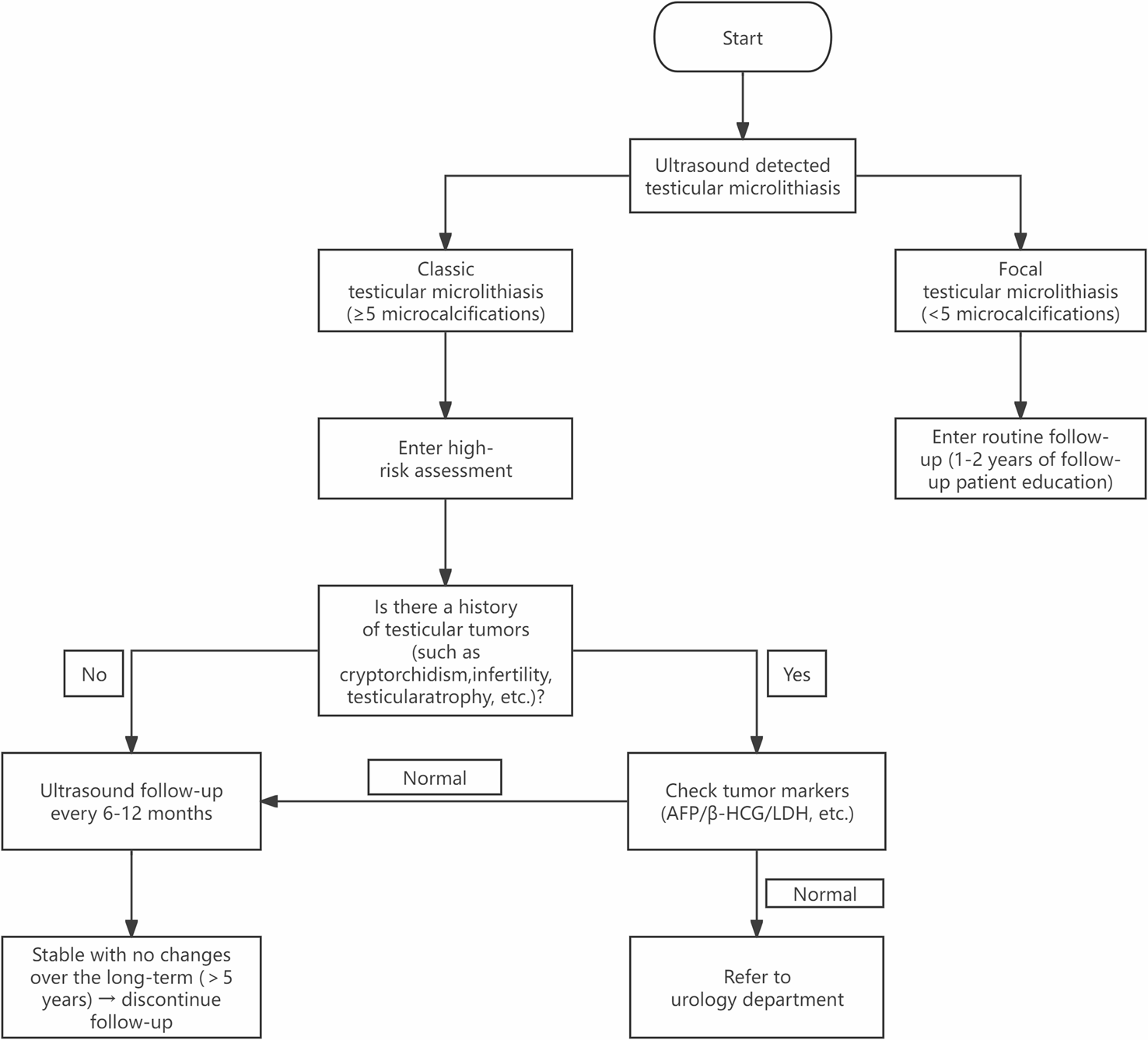

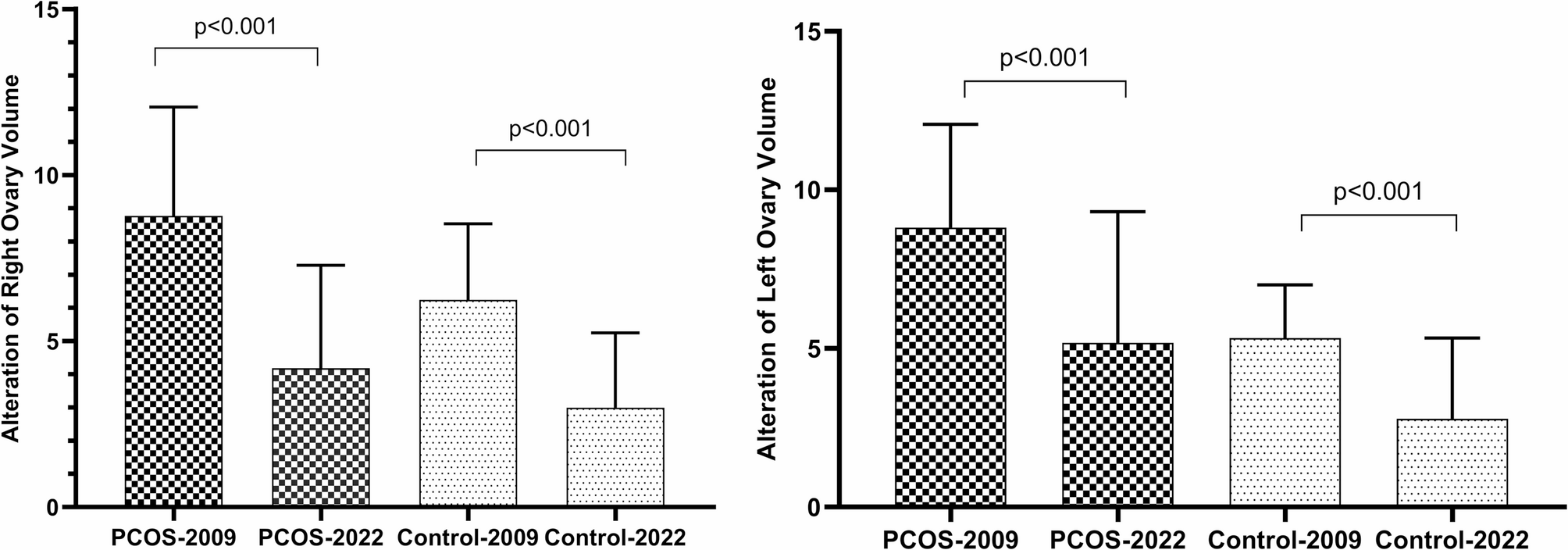





In order to investigate the potential impact of various TUBB8 variants on the in vitro maturation process of oocytes, we performed microinjections of WT and four mutant TUBB8 mRNA into mouse oocytes in vitro. The results revealed a significant reduction in the rate of first polar body extrusion in the p.A313V, p.G96R, p.C239W, and p.R251Q mutant TUBB8 groups compared to the WT group (Fig. 1A/B, 77.86% ± 2.80%, n = 152, vs. 47.08% ± 4.17%, n = 138, 56.93% ± 5.80%, n = 144, 39.97% ± 5.71%, n = 147, 36.14% ± 8.38%, n = 167, *P < 0.05; **P < 0.01). These findings suggested that TUBB8 variants led to a decrease in the extrusion rate of the polar body during the in vitro oocyte maturation process.
Fig. 1
The rate of extrusion of the first polar body in mouse oocytes with TUBB8 variants. (A) Representative images of in vitro matured oocytes. (B) Statistics of the rate of first polar body extrusion in mouse oocytes in vitro. Data are presented as mean ± standard deviation (mean ± SD), *P < 0.05; **P < 0.01
TUBB8 variants led to abnormal spindle morphology in mouse oocytesMicrotubule proteins are the main constituents of oocyte spindles, and TUBB8 is an important gene encoding microtubule protein. After microinjection of WT and four mutant TUBB8 mRNA into mouse oocytes in vitro, we found that oocytes in the p.A313V, p.G96R, p.C239W, and p.R251Q variant groups exhibited varying degrees of spindle abnormalities. In the A313V group, the spindle fibers showed no obvious abnormalities, while in the G96R group, the spindle fibers were disorganized, with uneven distribution and asymmetrical polarity. The C239W group showed dispersed spindle fibers, losing polarity, and the R251Q group exhibited a loss of normal spindle structure (Fig. 2A). Furthermore, we observed the presence of abnormal spindle types in all groups, such as spindles lacking proper structure, multiple spindle structures, multipolar spindles, short spindles, and asymmetric spindles (Fig. 2B). Statistical analysis revealed a significant increase in the percentage of abnormal spindles in the p.A313V, p.G96R, p.C239W, and p.R251Q mutant TUBB8 groups compared to the WT group (Figs. 2C and 24.67% ± 4.51%, n = 39, vs. 64.67% ± 9.50%, n = 51, 72.67% ± 10.69%, n = 45,61.67% ± 12.34%, n = 47, 73.00% ± 9.165, n = 49, **P < 0.01; ***P < 0.001.).
Fig. 2
TUBB8 variants lead to abnormal spindle morphology in mouse oocytes. (A) Representative images of spindle morphology in mouse oocytes at the metaphase II (MII) stage, with Flag indicating the signal of exogenous protein transduction, scale bar = 5 μm. (B) Abnormal spindle types in mouse oocytes at the MII stage, with Flag indicating the signal of exogenous protein transduction, and the white dashed box indicating the first polar body, scale bar = 5 μm. (C) Statistical analysis of the percentage of abnormal spindles in mouse oocytes. Data are presented as mean ± standard deviation (mean ± SD), **P < 0.01; ***P < 0.001
TUBB8 variants induced aberrant chromosome distribution in mouse oocytesTo further explore the impact of TUBB8 variants on chromosome distribution in oocyte spindles, we analyzed the chromosome plate structure in the spindles of each group of oocytes (Fig. 3A). The results showed that compared to the WT group, the p.A313V, p.G96R, p.C239W, and p.R251Q mutant TUBB8 groups exhibited a significant increase in the longitudinal axis width of the chromosome plate in the metaphase II (MII) oocyte spindles (Fig. 4A/B, 7.96 ± 2.02, n = 42, vs18.02 ± 4.19, n = 50, 24.44 ± 6.365, n = 47, 21.52 ± 6.68, n = 49, 20.81 ± 3.87, n = 50, *P < 0.05; **P < 0.01.). This suggested that the various TUBB8 variants affected the arrangement, distribution density, and dynamics of microtubule proteins in the spindle structure, leading to abnormal chromosome plate distances. This uneven distribution of chromosomes can further result in the occurrence of oocyte aneuploidy.
Fig. 3
Changes in mitochondrial distribution in mouse oocytes caused by TUBB8 variants (A) Mitochondrial distribution patterns in mouse oocytes during GV and MII stages, scale bar = 10 μm. (B) Statistical analysis of mitochondrial distribution proportions in mouse oocytes at the MII stage; data are presented as mean ± standard deviation (mean ± SD), *P<0.05; **P<0.01; ***P<0.001
Fig. 4
Abnormal chromosome distribution in mouse oocytes caused by TUBB8 variants. (A) Chromosome plate structure in the spindles of mouse oocytes at the metaphase II (MII) stage, scale bar = 5 μm. (B) Statistical analysis of chromosome plate width in the spindles of mouse oocytes at the MII stage. Data are presented as mean ± standard deviation (mean ± SD), *P < 0.05; **P < 0.01
TUBB8 variants did not affect the cortical polarity of mouse oocytesThe haploid oocyte generation process involves meiotic and asymmetric cell division. The polarity of oocytes determines the formation position and size of the two polar bodies, ensuring the production of haploid oocytes while retaining almost all cytoplasm within the oocyte for early embryonic development [21,22,23].The polarity of oocytes is crucial for proper meiotic division and spindle movement, and it is an important manifestation of oocyte maturation. Immunofluorescence staining of oocytes in different groups revealed that both wild-type (WT) and TUBB8 mutant oocytes at the metaphase II (MII) stage can form normal F-actin caps (Fig. 5), indicating that the TUBB8 variants do not affect the polarity of mature oocytes.
Fig. 5
Formation of the cortical actin cap in mouse oocytes. F-actin represents the cortical actin cap structure in oocytes, scale bar = 10 μm
TUBB8 variants disrupted mitochondrial distribution in mouse oocytesTwo distribution patterns of mitochondria are observed during oocyte maturation: clustered and uneven distribution, and uniform distribution (Fig. 3A). The overall movement and distribution pattern of mitochondria in oocytes is from the vicinity of the nucleus toward the cortex, and the uniform distribution of mitochondria represents a higher level of cytoplasmic maturation in oocytes. The distribution and rearrangement of mitochondria are mainly achieved through the pulling action of microtubule proteins. To investigate whether TUBB8 variants affect the rearrangement of organelles during oocyte maturation, we performed live staining of mitochondria in WT and TUBB8 variant oocytes. The results showed that the proportion of oocytes with uniformly distributed mitochondria at the metaphase II (MII) stage was significantly decreased in the p.A313V, p.G96R, p.C239W, and p.R251Q TUBB8 mutant groups compared to the WT group (Figs. 3B and B and 72.33 ± 2.51, n = 58, vs. 38.33 ± 10.40, n = 49, 26.67 ± 10.41, n = 50, 38.67 ± 5.69, n = 53, 21.00 ± 9.54, n = 55, *P<0.05; ** P<0.01;*** P<0.001). This indicates that the TUBB8 variants cause rearrangement and redistribution of mitochondria during oocyte maturation, leading to a decrease in cytoplasmic maturation of oocytes.
TUBB8 variants triggered an endoplasmic reticulum (ER) stress responseWe further examined the distribution of the endoplasmic reticulum (ER) and the calcium ion concentration in the cytoplasm of oocytes. The results revealed an increased content of ER (Fig. 6A/B, 16.75 ± 1.56, n = 45, vs. 28.71 ± 4.69, n = 53, 25.48 ± 4.67, n = 50, 26.73 ± 4.73, n = 52, 23.94 ± 5.80, n = 47, *P<0.05; ** P<0.01) and an elevated calcium ion concentration (Fig. 6A/C, 9.77 ± 2.43, n = 54, vs. 15.59 ± 2.90, n = 59, 18.02 ± 2.80, n = 52, 24.65 ± 6.68, n = 48, 20.61 ± 3.92, n = 51, *P<0.05; ** P<0.01), indicating impaired ER calcium storage and release capacity, leading to ER stress. These findings suggest an endoplasmic reticulum (ER) stress response, a reduced cytoplasmic maturation and compromised developmental potential of oocytes.
Fig. 6
TUBB8 variants induces endoplasmic reticulum (ER) stress response in mouse oocytes. (A) Fluorescence images showing the distribution of ER in oocyte cytoplasm; fluorescence images depicting the distribution of calcium ions in oocytes. Scale bar = 10 μm. (B) Analysis of ER fluorescence intensity in oocytes. (C) Analysis of calcium ion fluorescence intensity in oocytes. Data are presented as mean ± standard deviation (mean ± SD), *P<0.05; ** P<0.01
Comments (0)