
Remember me
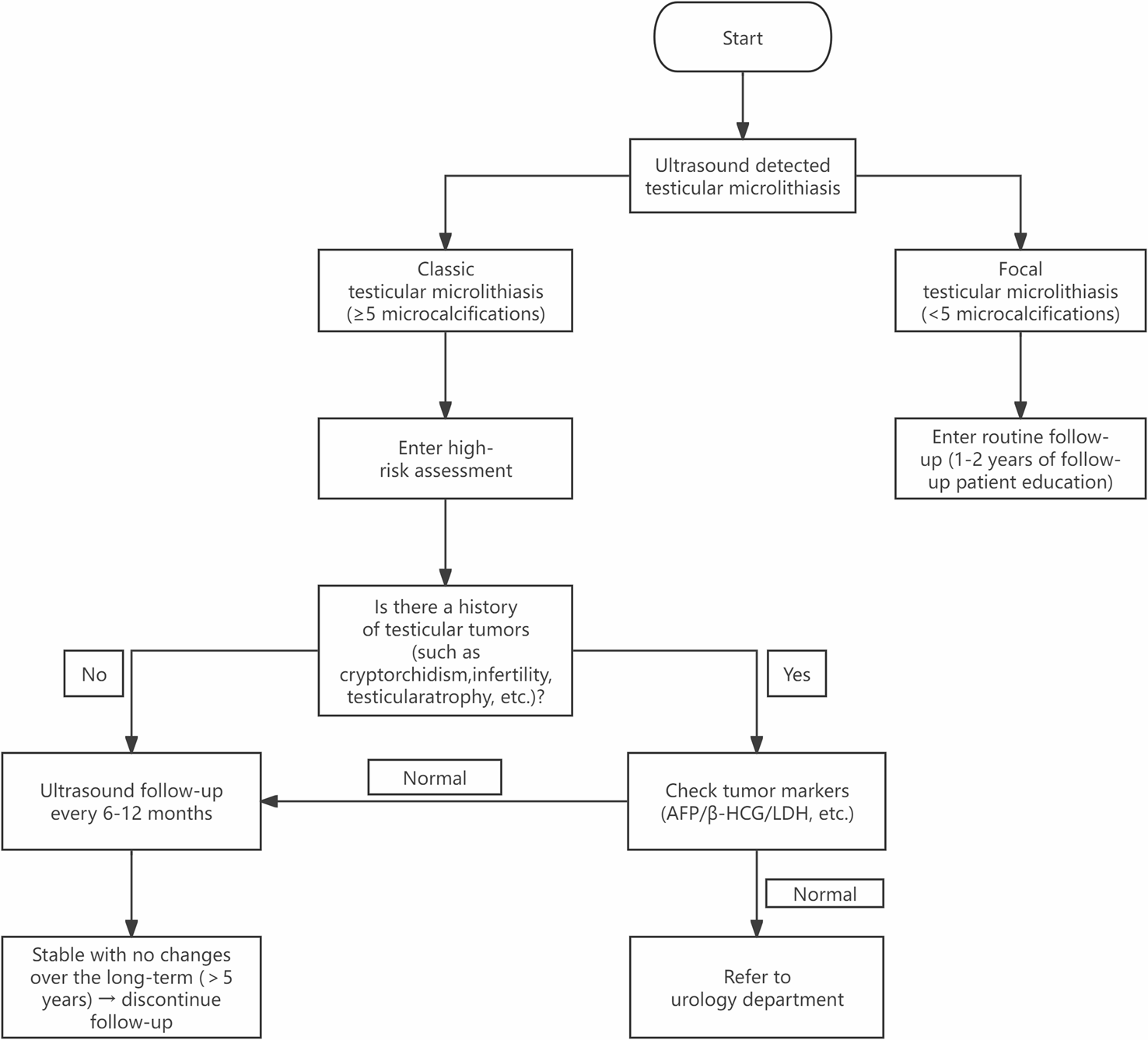






In this study, we selected the most widely used BMSCs in stem cell research to test the potency of Cargocytes in TD. First, we labeled BMSCs with Mitotracker red. Then, we enucleated BMSCs using Cytochalasin B and Ficoll density gradient ultracentrifugation (Fig. 1A). The predominant components from top to bottom were cell debris, enucleated BMSCs and nucleoplasts (Fig. S1A), respectively. Next, we found that Cargocytes were significantly smaller than BMSCs and readily attached to tissue culture plates with well-organized cytoskeletal structure (Fig. 1B, C). Moreover, Cargocytes contained crucial subcellular organelles including mitochondria, ER, and lysosomes (Fig. 1D, E). The co-staining of Mitotracker and actin indicated that enucleated BMSCs had a complete cytoskeleton so that mitochondria could be well preserved as BMSCs (Fig. S1B). Hereafter, to assess the mitochondria of Cargocytes, we used TMRE kit to analyze the mitochondrial membrane potential. The flow cytometry results indicated that Cargocytes had normal mitochondrial membrane potential (Fig. 1F, G). To examine whether enucleation affected the characteristics of BMSCs, we used flow cytometry to analyze the surface markers of BMSCs. And we found that Cargocytes expressed the same panel of surface markers of BMSCs, including CD29, CD90, and CD105, and did not express CD34 or CD45, indicating that Cargocytes maintain the phenotype of BMSCs (Fig. 1H).
Fig. 1
Enucleated BMSCs retained important features. A Schematic of workflow for preparation of enucleated BMSCs (Cargocytes). B Espial of organelles through staining with mitochondrial and endoplasmic reticulum fluorescent probes. DAPI (blue) indicated the presence of the nucleus. The lysosome was stained by LAMP1. Scale bar, 20 μm. C Quantitative analysis of the MFI of Mitotracker, ER tracker and LAMP1 in B. n = 3 biological repeats for each group; unpaired t test. Two-sided comparison; All data are Mean ± SEM. D The expression of cytoskeletal markers α-tubulin and actin in BMSCs and enucleated BMSCs. Nucleus was detected by DAPI staining (blue). Scale bar, 20 μm. E Quantitative analysis of the mean fluorescence intensity of α-tubulin and actin in D. n = 3 biological repeats for each group; unpaired t test. Two-sided comparison; Data are shown as the Mean ± SEM. F Mitochondrial membrane potential was analyzed using TMRE kit by flow cytometry. G Quantitative analysis of the MFI ratio of TMRE in F. n = 3 biological repeats for each group; unpaired t test. Two-sided comparison; Data are shown as the Mean ± SEM. H The expression levels of CD34, CD45, CD29, CD90 and CD105 were analyzed by flow cytometry
Cargocytes retain important cellular functionsAfter that, we investigated the ability of proliferation, migration and secretion of Cargocytes. We found that Cargocytes were viable for up to 72 h post-enucleation, as shown by the trypan blue dye exclusion assay (Fig. 2A, B). Consistent with previous studies [11, 14], both BMSCs and Cargocytes migrated towards the high concentration serum (Fig. 2C, D). Interestingly, Cargocytes and BMSCs had similar characteristic cup-shaped morphology by electron microscopy (Fig. 2E and S2), similar size distribution under DLS analysis (Fig. 2F). Moreover, Cargocytes had similar zeta potential as parental BMSCs (Fig. 2G), suggesting that Cargocytes might actively produce and release EVs. It is well known that MSCs have immunoregulatory abilities [8,9,10], we co-cultured activated Jurkat cells, the human acute T-cell leukemia cell line, with BMSCs or Cargocytes for 48 h. And then we detected the percentage of IL-10+ (anti-inflammatory cytokine) (Fig. 2H, I) and TNF-α+ (pro-inflammatory cytokine) Jurkat cells (Fig. 2J, K), and found that both BMSCs and Cargocytes suppressed the inflammatory reaction of activated Jurkat cells. Taken together, these results indicate that Cargocytes retain critical cell structures and functions, and therefore have potential for in vivo regulating immune response.
Fig. 2
Enucleated BMSC retained cellular functions. A The images of 24 h, 48 h, 72 h, and 96 h BMSCs and Cargocytes after trypan blue dye exclusion assay. Scale bar: 50 μm. B Graphs showed the population of viable BMSCs and Cargocytes. Data are shown as the Mean ± SEM; n = 3 biological replicates. C BMSCs and Cargocytes migrated in chambers towards FBS for 2 h. Representative images of BMSCs or Cargocytes that migrated to the underside of 8.0 μm porous filters were stained with Crystal Violet. Scale bar, 50 μm. D Bar graph represented the migrated cell number. Data are shown as the Mean ± SEM; n = 10 independent fields from 3 biological replicates. E Electron microscopy images of EVs from BMSCs and Cargocytes. Representative images out of 10 images obtained were shown. Arrowheads point to typical EVs. Scale bar, 200 nm. F Histograms showing the size of EVs from conditioned media of BMSCs or Cargocytes. G Bar graph showed the zeta potential of EVs measured by a Zetasizer Nano ZSE analyzer and analyzed by Zetasizer Software. H, J Jurkat cells were cultured with or without BMSCs or Cargocytes for 2 days, and their production of TNF- α and IL-10 was examined by flow cytometry. n = 3 biological replicates. I, K Bar graphs showed TNF- α+ and IL-10+ Jurkat cells after coculture with BMSCs and Cargocytes. Cumulative results are expressed as Mean ± SEM. (*P < 0.05, **P < 0.01, ***P < 0.001)
Cargocytes and BMSCs contribute to the number of interstitial cells in vivoMany studies have reported that transplanted MSCs and SLCs can recover testosterone production in vitro and in vivo [15]. However, these cells had heterogeneity and multi-directional differentiation potential. Based on the characteristics of Cargocytes, we considered if Cargocytes could restore LCs in TD rats. We established the EDS-treated model and injected Mitotracker red+ BMSCs or Cargocytes into the parenchyma of testes. At 0, 4, 8, 10, 12, and 14 days after EDS treatment, the serum and the testes were collected for analyses (Fig. 3A). Moreover, through immunofluorescence staining of the cell proliferation marker Ki67, it was found that the EDS + BMSCs group had more Ki67+ stromal cells than the EDS + Saline group, and the EDS + Cargocytes group had more Ki67+ cells than the EDS + BMSCs group 10 days after transplantation (Fig. 3B, C). The Hedgehog pathway has been reported to play an important role in the development of reproductive system [16] which may explain the increase of stromal cells. So, we sorted LHR+ cells from testes 10 days after implantation by FACS, and detected the expression of Hedgehog pathway ligands (Dhh, Shh, and Ihh), receptors (Smo and Ptch1), nuclear transcription factors (Gli1, Gli2, and Gli3), and target genes (Hhip and Hhat). In addition to Shh and Ihh, the expression of other genes significantly increased in the EDS + BMSCs group. Compared to the EDS + BMSCs group, the expression of these genes increased even more in the EDS + Cargocytes group (Fig. 3D). Furthermore, we detected protein levels of desert hedgehog (DHH), smoothened, frizzled class receptor (SMO), and GLI family zinc finger 1 (GLI1), which were consistent with mRNA changes and significantly increased in the EDS + BMSCs group and EDS + Cargocytes group (Fig. 3E, F). The above results indicate that BMSCs and Cargocytes may increase the quantity of stromal cells through the Hedgehog pathway in vivo.
Fig. 3
Transplanted BMSCs or Cargocytes contributed to the number of interstitial cells by Hedgehog pathway. A Schematic of the experimental procedure used for cell transplantation. B The proliferation of stromal cells of EDS-treated mice 10 days after implantation were stained by Ki67 (red) antibody. Nucleus was detected by DAPI staining (blue). Scale bar, 50 μm. C Quantitative analysis showing the number of Ki67+ cells per scope in the interstitial area of the testes. Data are shown as the Mean ± SEM; n = 6. D qRT-PCR analysis showed the expression of Hedgehog pathway in the LHR+ cells sorted from EDS-treated mice 10 days after implantation. Data are shown as the Mean ± SEM; n = 6. E Western blot analysis of the protein levels of DHH, SMO and GLI1 in the LHR+ cells sorted from EDS-treated mice 10 days after implantation. F Quantitative analysis showing the expression of proteins. Data are shown as the Mean ± SEM. (*P < 0.05, **P < 0.01, ***P < 0.001)
Transplanted Cargocytes and BMSCs can restore testosterone productionLCs are the primary source of testosterone in the interstitial compartment of testis [17]. We found that the mRNA level of LC markers LhcgR, Cyp11a1, and Cyp17a1 significantly increased 10 days after transplantation. Importantly, the testes of EDS + Cargocytes group showed higher expression levels of LC markers than the EDS + BMSCs group (Fig. 4A). Together, these data suggested that transplanted Cargocytes might induce more LCs than BMSCs. After that, we detected the levels of serum testosterone at different time points. Consistent with previous observation [18], the concentration of serum testosterone was decreased 4 days after EDS treatment, indicating that EDS specifically eliminated the testosterone-producing LCs (Fig. 4B). The serum testosterone levels of rats with BMSCs or Cargocytes transplantation were increased significantly compared with the EDS-treated rats with saline injection. Moreover, the EDS-treated rats produced more testosterone after Cargocytes injection (Fig. 4B). Besides, immunohistochemical analysis showed that the expression of steroidogenic enzymes 3β-HSD (Fig. 4C and 4D) and StAR (Fig. 4E, F) was increased 10 days after implantation. Notably, the EDS + Cargocytes group expressed higher levels of 3β-HSD and StAR than the EDS + BMSCs group. Taken together, these results demonstrate that Cargocytes are capable of restoring the number of LCs and recovering testosterone production.
Fig. 4
Transplanted BMSCs or Cargocytes increased the number of LCs and testosterone production. A qRT-PCR analysis showed the expression of LhcgR, Cyp11a1 and Cyp17a1 in the testes of EDS-treated rats 10 days after implantation. B The serum testosterone concentration was measured at the indicated time points in each animal. Data are shown as the Mean ± SEM; n = 6. C Immunofluorescence staining showed the accumulation of 3β-HSD+ cells (green) in the interstitial area of the testes of EDS-treated rats 10 days after implantation. Scale bar, 50 μm. D Quantitative analysis showing the number of 3β-HSD+ cells per scope in the testes of EDS + Saline, EDS + BMSCs and EDS + Cargocytes groups. Data are shown as the Mean ± SEM; n = 6. E Immunofluorescence staining showed the accumulation of StAR+ cells (magenta) in the interstitial area of the testes of EDS-treated rats 10 days after implantation. Scale bar, 50 nm. F Quantitative analysis showing the number of StAR+ cells per scope in the testes of EDS + Saline, EDS + BMSCs and EDS + Cargocytes groups. Data are shown as the Mean ± SEM; n = 6. (*P < 0.05, **P < 0.01, ***P < 0.001)
Transplanted Cargocytes and BMSCs offer potential benefits for spermatogenesisThe testes are the only place for sperm production, and in males, the hypothalamic pituitary testicular axis affects sperm production by regulating testosterone [14]. EDS reduces the elimination of LCs, thereby affecting the spermatogenic function [15]. To further investigate whether BMSCs and Cargocytes could restore spermatogenic function in vivo, we examined the number of spermatogonia in the seminiferous tubules 10 days after transplantation. DAZL+ cells almost disappeared after EDS + Saline treatment, but EDS rats transplanted with BMSCs significantly restored spermatogonia. EDS rats injected with Cargocytes had the highest number of spermatogonia (Fig. 5A, B). Spermatogonia undergo meiosis to form secondary spermatocytes, which further differentiate into mature sperm. This indicated that the process of sperm generation was accompanied by meiosis. Therefore, we observed the number of SYCP3+ cells representing meiosis in the seminiferous tubules 10 days after transplantation. We found that only a few cells in the EDS + Saline group were undergoing meiosis, while Cargocytes were able to increase the number of meiotic cells more effectively than BMSCs (Fig. 5C, D). Correspondingly, the number of CREM+ sperm cells showed a similar trend of change with DAZL and SYCP3 (Fig. 5E, F). The above results indicate that BMSCs and Cargocytes can promote the production of testosterone, and restore spermatogenic function.
Fig. 5
Transplanted BMSCs or Cargocytes promoted spermatogenesis. A The spermatogonium were detected by immunofluorescence staining with anti-DAZL (magenta) antibody. B Quantitative analysis showing the percentage of DAZL+ cells in seminiferous tubules of testes. Data are shown as the Mean ± SEM; n = 6. C The meiotic spermatocytes were observed by immunofluorescence staining with anti-SYCP3 (green) antibody. Nucleus was detected by DAPI staining (blue). The SYCP3+ cells were located at the pachytene stage in each testis. D Quantitative analysis showing the percentage of SYCP3+ cells in seminiferous tubules of testes. Data are shown as the Mean ± SEM; n = 6. E Representative images showed CREM+ cells (red) in seminiferous tubules of testes 10 days after transplantation. Scale bar, 50 µm. F Quantitative analysis showing the percentage of CREM+ cells in seminiferous tubules of testes. Data are shown as the Mean ± SEM; n = 6. (*P < 0.05, **P < 0.01, ***P < 0.001)
Enucleated BMSCs promote macrophages M2 polarization in vivoTo examine the role of Cargocytes in the benefits of testosterone production and spermatogenesis in vivo, we stained the testes sections with actin 4 days after transplantation. Fluorescence results showed that at this time, BMSCs still remained intact cytoskeletal structure, while the skeleton of Cargocytes had been broken (Fig. 6A). It is well known that macrophages can actively phagocytose broken and apoptotic cells and remodel the immune microenvironment [19, 20]. Furthermore, we sought to confirm whether BMSCs and Cargocytes could regulate the interstitial testicular macrophages (TMs) in vivo. We found that F4/80+ macrophages appeared in the EDS + Saline testes 4 days after transplantation, while the testicular interstitium of the EDS + BMSCs group and the EDS + Cargocytes group contained more macrophages (Fig. S3A, B), which suggested that interstitial TMs might aggregate in response to the injury of LCs. Meanwhile, we found that F4/80+ macrophages colocalized with Cargocytes, but not BMSCs (Fig. 6B). Collectively, we found that CD86+ M1 macrophages were decreased significantly after transplantation, while CD163+ M2 macrophages were increased (Fig. 6C, D). Notably, there were fewer CD86+ macrophages and more CD163+ macrophages in the EDS + Cargocytes group than in the EDS + BMSCs group. This phenomenon was aggravated with time (Fig. S3C-F). The anti-inflammatory effect of Cargocytes was better than that of BMSCs. Moreover, considering the injury of LCs might induce the production of reactive oxygen species (ROS), we stained the testes sections with DHE 10 days after transplantation, and found that the levels of ROS in the EDS + Saline group was very high. Cell transplantation could reduce the ROS, and the transplantation of Cargocytes could obtain better effect than BMSCs (Fig. 6E, F). In addition, the expression of Il-6 and Tnf-α was decreased, while the expression of Il-10 and Tgf-β was increased in the cell transplantation groups (Fig. 6G). The EDS + Cargocytes group expressed lower CD86 and nitric oxide synthase 2 (Nos2), the markers of M1 macrophages, and higher CD206 and arginase 1 (Arg1) (the markers of M2 macrophages) 10 days after transplantation (Fig. 6H). The above results indicate that after the transplantation of Cargocytes, interstitial TMs polarize from M1 to M2 and remodel the immune microenvironment and redox microenvironment.
Fig. 6
Transplanted Mitotracker red+ BMSCs or Cargocytes promoted macrophage polarization. A Immunofluorescence staining showed the colocalization of cells positive for Mitotracker (red) and F4/80 (green) in the testes of EDS-treated rats 4 days after implantation. Scale bar, 20 μm. B Immunofluorescence double-staining results of Mitotracker (red) and actin (green) in the testes of EDS-treated rats 4 days after implantation. Nucleus was detected by DAPI staining (blue). Scale bar, 20 μm. C Immunofluorescence staining showed the accumulation of CD86+ cells (green) and CD163+ cells (magenta) in the testes of EDS-treated rats 4 days after implantation. Scale bar, 50 μm. D Quantitative analysis showing the number of CD86+ cells (green) and CD163+ cells (magenta). E Fluorescent staining of testis slices with DHE to detect ROS. F Quantitative analysis showing the MFI of DHE. Data are shown as the Mean ± SEM; n = 6. G qRT-PCR analysis showed the expression of Il6, Tnf-α, Il10 and Tgf- β 10 days after transplantation. H qRT-PCR analysis showed the expression of CD86, Nos2, CD206 and Arg1 10 days after implantation. (*P < 0.05, **P < 0.01, ***P < 0.001)
Comments (0)