
Remember me


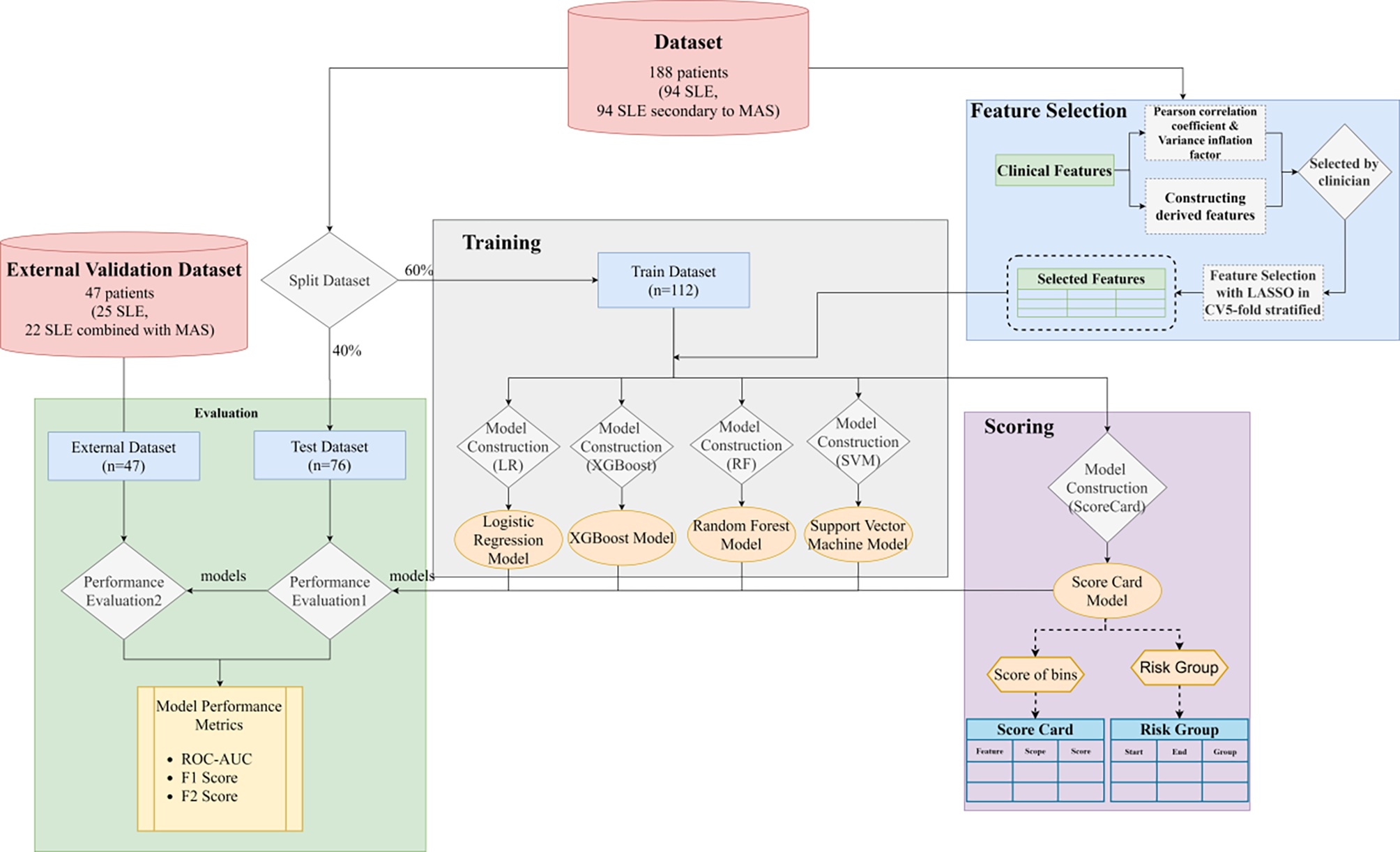
First, we investigated the frequency of Th1-like Tregs in the peripheral blood of SLE patients and HDs. CD4+FOXP3+ total Tregs, CD4+CXCR3+FOXP3+Th1-like Tregs, and CD4+CXCR3−FOXP3+ non-Th1-like Tregs were identified using flow cytometry. Although the frequency of CD4+FOXP3+ total Tregs was unchanged in SLE patients, the frequency of CD4+CXCR3+FOXP3+Th1-like Tregs, but not non-Th1-like Tregs, significantly increased in SLE patients compared to the HDs (Fig. 1A). We also found that the frequency of CD4+T-bet+FOXP3+Th1-like Tregs, but not CD4+T-bet−FOXP3+ non-Th1-like Tregs, was significantly increased in SLE patients compared to the HDs (Fig. 1B). In addition, we measured clinical immune indicators and SLE disease activity index (SLEDAI) in the SLE patients to evaluate disease activity and severity. A positive correlation was observed between the frequency of Th1-like Tregs and total immunoglobulin G (IgG) levels (Fig. 1C), suggesting that Th1-like Tregs might contribute to the SLE autoimmune response.
Fig. 1
Th1-like Treg cells increased, accompanied by low expression of TIGIT in SLE patients. A, C–E PBMCs isolated from the peripheral blood of 33 SLE patients and 15 HDs shown in Supplementary Table S1 were analyzed using flow cytometry without incubation. A Frequencies of Foxp3+Tregs, CXCR3+Foxp3+Th1-like Tregs, and CXCR3−Foxp3+non-Th1-like Tregs in CD4+ T cells. B PBMCs isolated from the peripheral blood of 16 SLE patients and 10 HDs shown in Supplementary Table S2 were analyzed using flow cytometry without incubation. Representative flow cytometry plots and bar graphs showing the frequencies of Foxp3+Tregs, T-bet+Foxp3+Th1-like Tregs, and T-bet−Foxp3+non-Th1-like Tregs in CD4+ T cells. C Correlations of levels of IgG and SLEDAI with the proportion of Foxp3+Tregs from SLE patients. D The proportion of TIGIT+ cells in CD4+Foxp3+Tregs. E Correlations of levels of C3 and C4 with the proportion of TIGIT+CD4+Foxp3+Tregs from SLE patients. ns, not significant, *P < 0.05, **P < 0.01
We assessed TIGIT expression in the Tregs and found that its expression was decreased in the CD4+FOXP3+ total Tregs of SLE patients (Fig. 1D). We also evaluated the correlation between disease severity and TIGIT expression in Treg cells of SLE patients and found that complement 3 (C3) and C4 were negatively correlated with TIGIT expression in the Tregs (Fig. 1E), suggesting that a higher proportion of Tregs exhibited the Th1 phenotype in SLE patients compared to HDs, with low TIGIT expression and disrupted immune suppression function, which is related to the severity of SLE disease.
IFN-α-mediated CD4+T cells are dependent on STAT1 signaling in out-of-sequence conditions and STAT4 signaling under in-sequence conditionsTo clarify the effect of IFN-α on Treg differentiation, in vitro cultures were established, simulating the sequential order of the activation of IFNAR and TCR signaling. CD4+T cells were isolated from HD PBMCs and stimulated with anti-CD3 and anti-CD28 antibodies to activate TCR signaling. Time-gradient experiments indicated that the increasing STAT1 phosphorylation levels (pSTAT1) induced by TCR stimulation reached stable levels after 48 h, indicating full activation of TCR signaling in CD4+T cells (Fig. 2A). Moreover, pSTAT1, pSTAT3, pSTAT4, and pSTAT5 levels in CD4+T cells were all elevated to varying degrees after stimulation with exogenous IFN-α in the absence of TCR signaling, with the most significant increase observed with pSTAT1. IFN-α elevated and stabilized pSTAT1 levels within 20 min of exposure of CD4+T cells without TCR stimulation (Fig. 2B), which was concentration independent (Fig. 2C). In contrast, 48 h after complete activation of TCR, pSTAT4 levels, not pSTAT1 levels, in CD4+T cells were significantly enhanced by the subsequent addition of exogenous IFN-α (Fig. 2D).
Fig. 2
The different order in which IFN-α and TCR activate CD4+T cells can lead to the activation of different STAT pathways. CD4+T cells purified from PBMCs of HDs were stimulated at different time points, then analyzed using phosflow technology. A Percentages of pSTAT1+ cells and pSTAT4+ cells after TCR stimulated for 20 min, 24 h, 48 h, or 120 h. B, C Percentages and MFI of pSTAT1+ cells, STAT3+ cells, STAT4+ cells, and pSTAT5+ cells under the 20-min stimulation of IFN-α with specified concentration without TCR. D After 48 h of TCR-stimulation, percentages of pSTAT1+cells, STAT3+cells, STAT4+cells, and pSTAT5+cells were measured after further stimulation with or without IFN-α (20 ng/ml) for 20 min. E Schematic diagram of the setting in-sequence or out-of-sequence for culture conditions. F Percentages of pSTAT1+ cells, STAT3+ cells, STAT4+ cells, and pSTAT5+ cells after stimulation under TCR, in-sequence or out-of-sequence condition for 5 days. G After 5 days of TCR-stimulation, percentages of pSTAT1+cells, STAT3+cells, STAT4+cells, and pSTAT5.+cells were measured after further stimulation with or without IFN-α for 20 min. Data are mean ± SEM of three independent experiments using different donors. un-stimul, un-stimulation; min, minutes; h, hours; d, days. *P < 0.05
Furthermore, we established out-of-sequence culture conditions by the addition of IFN-α to inactivated CD4+T cells 20 min before TCR stimulation and in-sequence culture conditions by adding IFN-α to CD4+T cells 48 h after TCR stimulation (Fig. 2E). To verify the long-term effects of the stimuli, CD4+ T cells were cultured for 5 days under only TCR activating, out-of-sequence, or in-sequence culture conditions. Under all conditions, pSTAT4 levels were relatively high after 5 days. Notably, pSTAT4 levels were significantly enhanced under in-sequence conditions compared to the other treatments (Fig. 2F). When cells cultured under only TCR stimulation for 5 days were subsequently stimulated by IFN-α for 20 min, pSTAT4, but not pSTAT1, was enhanced. These data suggested that the effect of IFN-α on CD4+ T cells was dependent on the STAT1 pathway under out-of-sequence conditions but dependent on the STAT4 pathway under in-sequence conditions.
IFN-α promotes Th1-like Treg transformation under in-sequence conditionsWe investigated whether the differentiation of Th1-like Tregs was sensitive to the sequential effects of IFN-α. CD4+CD25+Tregs were purified and cultured under only TCR stimulating, out-of-sequence, or in-sequence conditions for 5 days. We found that the differentiation of CD4+CXCR3+Foxp3+ Th1-like Tregs increased under in-sequence conditions, whereas they decreased under out-of-sequence conditions (Fig. 3A). The same results were observed when defining Th1-like Tregs using T-bet and Foxp3 (Fig. 3B). CXCR3 and T-bet expression levels in the Tregs were increased by IFN-α under in-sequence conditions but decreased under out-of-sequence conditions, whereas Foxp3 and CD25 expression levels were not significantly affected (Fig. 3C). In addition, we found that TIGIT expression in the Tregs was reduced under in-sequence conditions (Fig. 3D). Together, our findings suggested that IFN-α could promote the transformation of Tregs into Th1-like Tregs by decreasing TIGIT expression under in-sequence conditions.
Fig. 3
Th1-like Tregs were increased by IFN-α under in-sequence condition. CD4+CD25+Tregs purified from PBMCs of SLE patients were stimulated under TCR, out-of-sequence or in-sequence condition for 5 days, then analyzed using flow cytometry. A Percentages of CXCR3+Th1 cells, Foxp3+Tregs, and T-bet+Foxp3+Th1-like Tregs. B Percentages of T-bet+Th1 cells and T-bet+Foxp3+Th1-like Tregs. C Levels of T-bet, CXCR3, Foxp3, and CD25 expressed in MFI. D Percentages of TIGIT+ cells in Foxp3+Tregs. Data are mean ± SEM of five independent experiments using different donors. *P < 0.05, **P < 0.01
IFN-α-induced Th1-like Tregs exhibit immunosuppressive dysfunctionWe also investigated whether IFN-α induction affected Treg-mediated immunosuppression. TIGIT agonist was used to activate TIGIT in Tregs. Interestingly, TIGIT activation reversed the conversion of Th1-like Tregs induced by IFN-α under in-sequence conditions (Fig. 4A, B). We further examined cytokine levels in the cell culture supernatant and found decreased levels of the suppressive cytokine IL-10 and increased secretion of the Th1-related proinflammatory cytokine IFN-γ under in-sequence conditions. TIGIT activation reversed those changes (Fig. 4C). To assess the suppressive effect of Treg cells on B-cell differentiation, IFN-α-stimulated Tregs were pretreated with TIGIT agonist and then co-cultured with B cells. TIGIT activation rescued the Th1-like Treg-mediated increase in plasmablasts (Fig. 4D). Therefore, TIGIT activation restored the immunosuppressive function of Th1-like Tregs reduced by IFN-α.
Fig. 4
Impaired immunosuppressive function of Th1-like Tregs can be restored by reactivating TIGIT. CD4+CD25+Tregs purified from PBMCs of SLE patients were stimulated under TCR or in-sequence condition with or without TIGIT agonists for 5 days. A Percentages of CXCR3+Foxp3+Th1-like Tregs were analyzed using flow cytometry. B Percentages of T-bet+Foxp3+Th1-like Tregs were analyzed using flow cytometry. C Levels of IL-10 and IFN-γin cell culture supernatant were measured using ELISA. D CD25+Tregs were purified from cultured CD4+T cells, then co-cultured with CD19+B cells. Percentages of CD27+CD38.+plasmablasts were analyzed using flow cytometry. Data are mean ± SEM of five independent experiments using different donors. *P < 0.05, **P < 0.01
STAT4 is essential for IFN-α-mediated differentiation of Th1-like TregsWe investigated whether the effect of IFN-α on Th1-like Treg differentiation was dependent on the STAT4 pathway for the in-sequence effect. CD4+T cells were pretreated with IFN-α and siRNA for 48 h before stimulating TCR signaling (i.e., out-of-sequence conditions). STAT1 and STAT4 mRNA expression levels were effectively silenced by siRNA (Fig. 5A). In the absence of STAT1, proliferation and differentiation of CD4+ T cells failed. STAT4 knockdown marginally aggravated the IFN-α inhibitory effect on the conversion of CXCR3+Foxp3+Th1-like Tregs under out-of-sequence conditions (Fig. 5B). Under in-sequence conditions, silencing of STAT1 increased pSTAT4, and STAT4 knockdown increased pSTAT1 (Fig. 5C). STAT4 knockdown, but not that of STAT1, abolished the expression of T-bet and CXCR3 induced in Foxp3+Tregs (Fig. 5D, E). These results indicated that IFN-α improved Th1-like Tregs under in-sequence conditions in a STAT4-dependent manner.
Fig. 5
IFN-α improved Th1-like Tregs through STAT4 pathway in-sequence condition. CD4+ T cells were purified from PBMCs of HDs. A Relative messenger RNA expression of STAT1 and STAT4 in transfected CD4+ T cells was evaluated by quantitative PCR. B Knocking out STAT4 before exposing to specified conditions. Bar graphs showing the percentages of CXCR3+Foxp3+Th1-like Tregs analyzed using flow cytometry. C–E CD4+ T cells were activated by TCR before incubated under specified conditions and analyzed using flow cytometry. C Percentages of pSTAT1+ cells, and STAT4+ cells in Foxp3+Tregs. D Percentages of CXCR3+ cells and T-bet+ cells. E MFI of CXCR3 and T-bet in Foxp3+Tregs. Data are mean ± SEM of three independent experiments using different donors. *P < 0.05, **P < 0.01, **P < 0.001
Activating TIGIT restores the dysfunction of Tregs by specifically regulating the STAT4 pathway under in-sequence conditionsRecent studies showed that TIGIT can repress the Akt/mTOR pathway [25]. We measured the phosphorylation levels of Akt (pAkt) and mTOR (pmTOR) in Tregs and found that both pAkt and pmTOR levels were elevated under in-sequence conditions, but the addition of the TIGIT agonist suppressed these elevated levels (Fig. 4A). Under in-sequence conditions, activating TIGIT significantly reduced pSTAT4 levels induced by IFN-α but did not affect the pSTAT1 and pSTAT4 levels caused by TCR activation (Fig. 6B). To verify the effect of TIGIT upstream of TCR, cells were pretreated by TIGIT agonists before TCR activation. The results showed that pre-activation of TIGIT reduced both pSTAT1 and pSTAT4 levels (Fig. 6C) and weakened the differentiation of Th1-like Tregs (Fig. 6D).
Fig. 6
TIGIT can reverse Treg cell function and phenotype. CD4+ T cells purified from PBMCs of HDs were stimulated under specified conditions for 5 days and analyzed using flow cytometry. A Percentages of pAkt+ cells and mTOR + cells in Foxp3+Tregs. B Percentages of pSTAT1+ cells and STAT4+ cells in Foxp3+Tregs. C, D CD4+ T cells were pretreated using TIGIT agonists before exposing to TCR. C Percentages of pSTAT1+ cells, STAT4+ cells in Foxp3+Tregs. D Percentages of CXCR3+Foxp3+Th1-like Tregs. Data are mean ± SEM of seven independent experiments using different donors. *P < 0.05, **P < 0.01, **P < 0.001
The in-sequence effect of IFN-α dominates in SLE patients, thus exerting the effect of promoting Th1-like Treg depending on pSTAT4Finally, we investigated whether IFN-α-mediated in-sequence effects caused the increase in Th1-like Tregs in SLE patients. The results showed that the expression levels of CD69, PD-1, and CD25 in the Tregs were all higher in SLE patients than in HDs (Fig. 7A). Without stimulation, the levels of pSTAT1 and pSTAT4 in the CD4+CD25+Tregs from SLE patients were higher than in HDs. IFN-α stimulation induced a more significant increase in pSTAT4 levels in the SLE patients (Fig. 7B). To explore the therapeutic effects of TIGIT on SLE, Tregs were treated with a TIGIT agonist and cultured under TCR stimulating conditions for 5 days. We found that the increase in the proportion of Th1-like Tregs was inhibited by the TIGIT agonist (Fig. 7C). In conclusion, our study showed that in SLE patients, Tregs were in a high TCR-activated state, and IFN-α could induce their differentiation into Th1-like Tregs through a STAT4-dependent in-sequence process, which could be reversed by TIGIT activation.
Fig. 7
IFN-α mediated highly activated T cells following in-sequential effects in SLE. A PBMCs isolated from the peripheral blood of 16 SLE patients and 10 HDs shown in Supplementary Table S2 were analyzed using flow cytometry without incubation. Bar graphs showing the frequencies of CD69+ cells, PD-1+ cells, and CD25+ cells in CD4+ T cells. B Whole blood of patients with SLE was collected, stained directly or after the stimulation by IFN-α for 20 min. Bar graphs showing the MFI of pSTAT1 and pSTAT4 in Foxp3+Tregs analyzed using flow cytometry. C CD4+CD25+Tregs purified from ten PBMCs of SLE patients were stimulated under specified conditions for 5 days and analyzed using flow cytometry. Representative flow cytometry plots and bar graphs showing the frequencies of CXCR3+Foxp3+Th1-like Tregs in CD4+ T cells
Comments (0)