
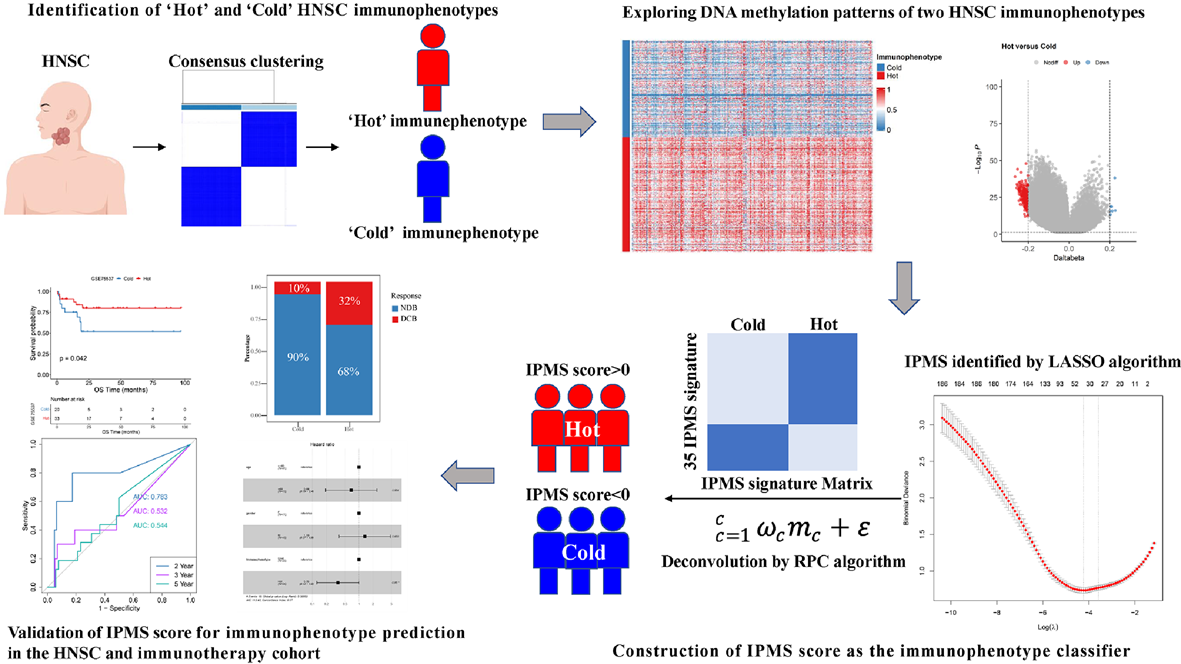
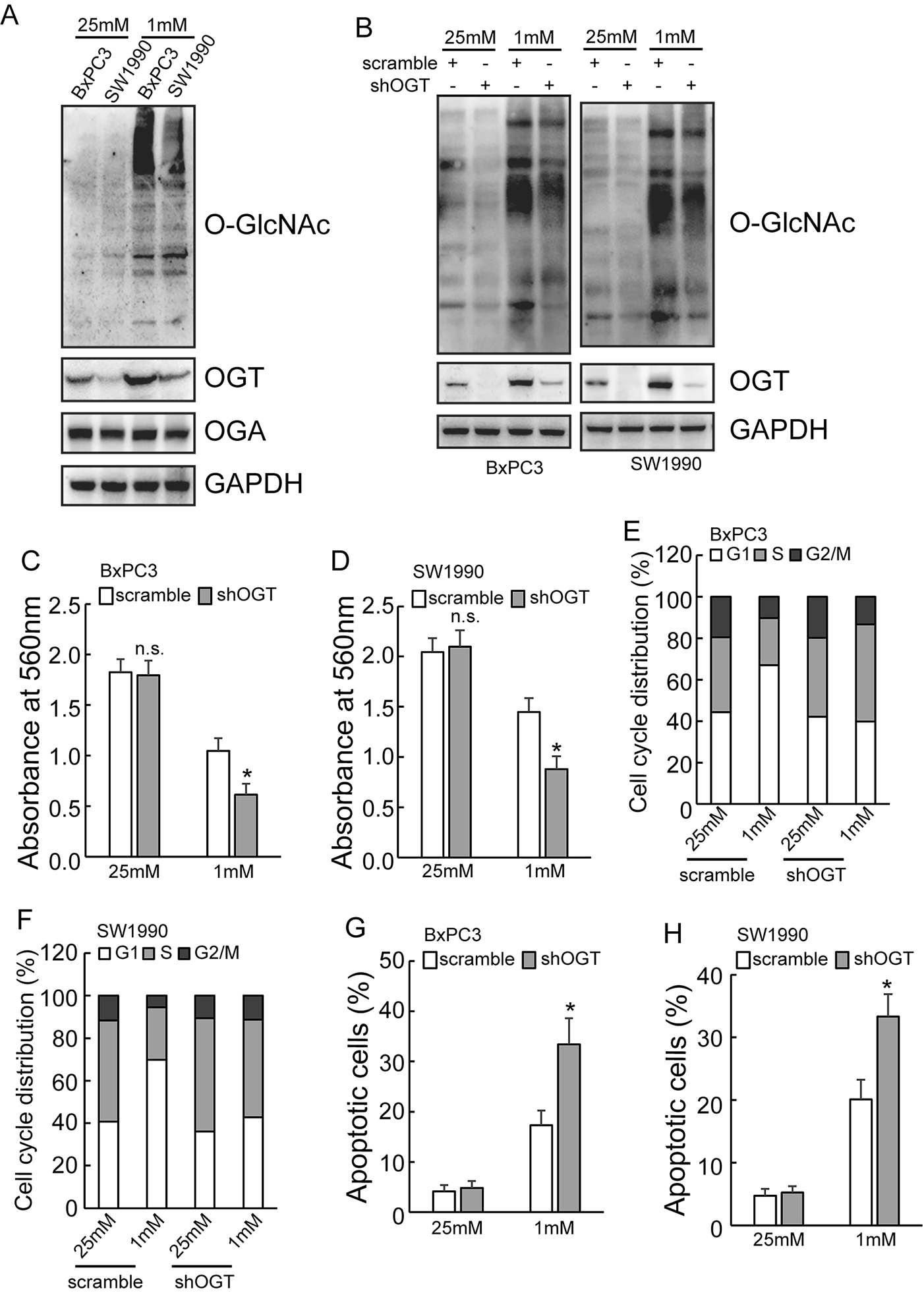
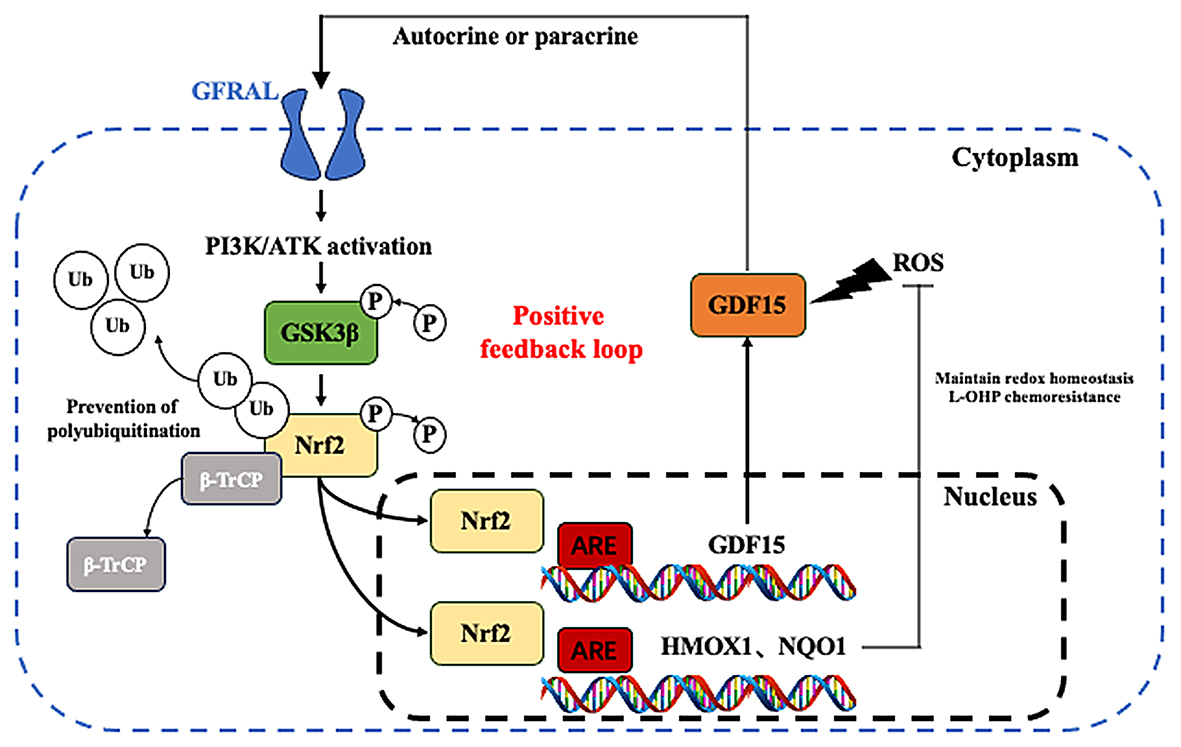
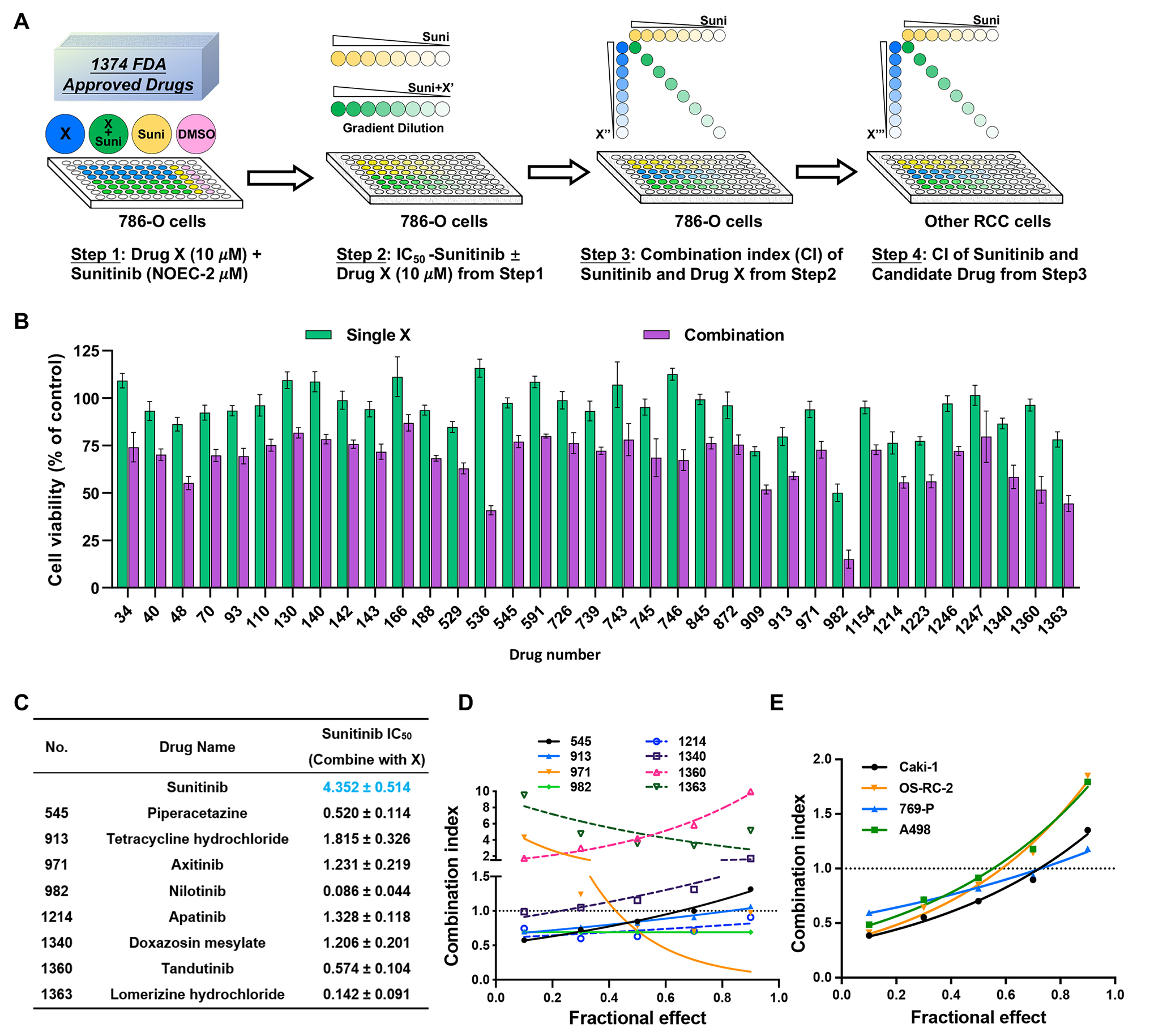
記住我



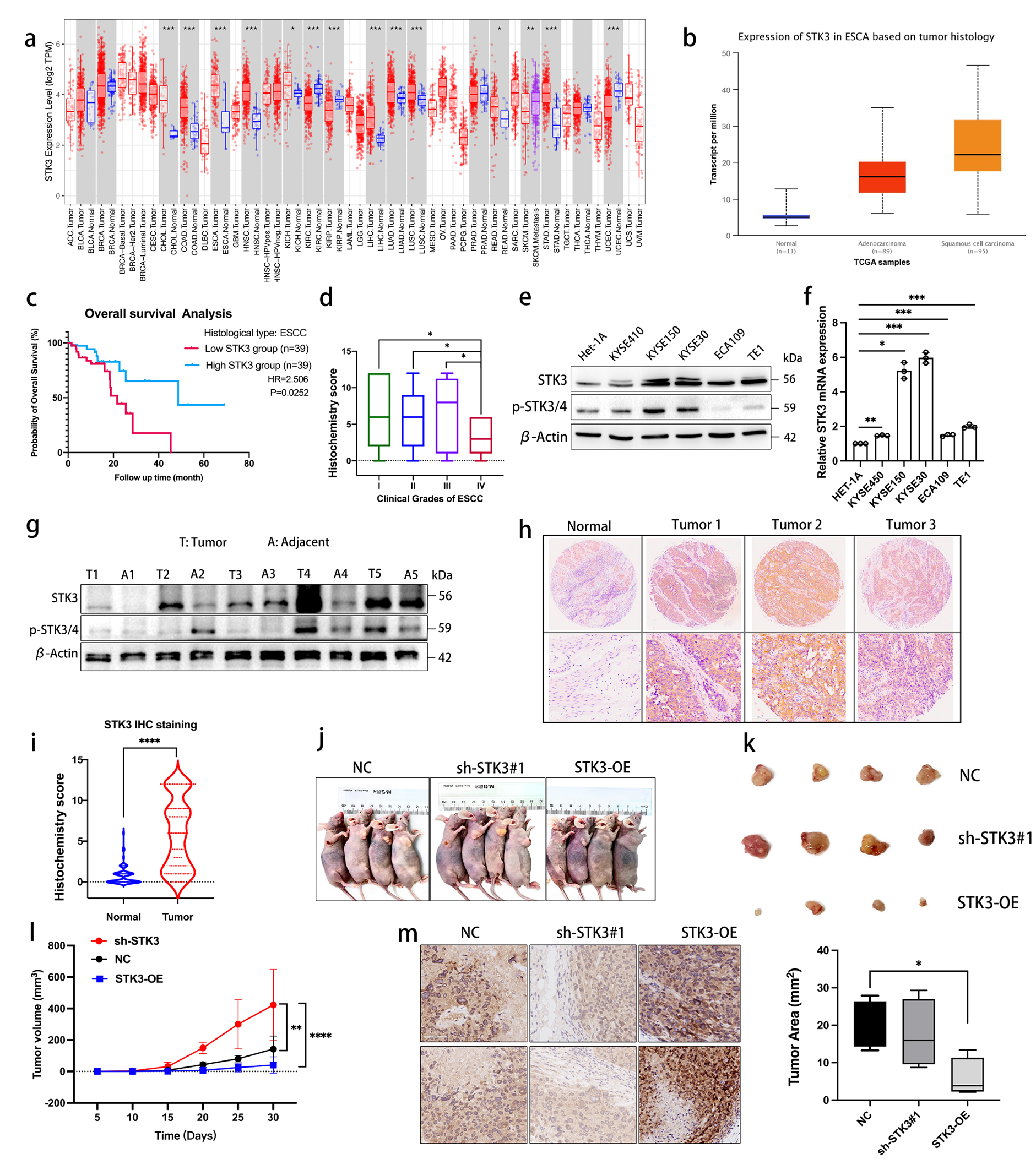
CT45A1 expression was investigated in cervical cancer patients. Immunofluorescence (IF) staining revealed that CT45A1 was overexpressed in the tumor tissues of cervical cancer patients, but not in the paired paracancerous tissues (Supplementary Fig. S1A). Immunohistochemical (IHC) staining revealed that the CT45A1 level in the tumor tissues obtained from 119 cervical cancer patients (Supplementary Table S1) was markedly higher than that in the paired paracancerous tissues (Fig. 1A, B, p < 0.001), with 88% specificity and 62% sensitivity without classification of cancer stage. Among the 20 benign uterine myoma patients, IHC staining showed that 16 patients did not express CT45A1, 3 patients barely expressed CT45A1, only 1 patient expressed CT45A1 at a moderate level (Fig. 1C, D), indicating that there was virtually no expression of CT45A1 in most of the benign uterine myoma patients. The CT45A1 level of patients with early-stage cervical cancer (I–II) was significantly higher than that of benign uterine myoma patients (p < 0.001) (Fig. 1D). The CT45A1 level of patients with advanced-stage cervical cancer (III-IV) was much higher than that of benign uterine myoma patients (Fig. 1D, p < 0.001). Strikingly, the specificity and sensitivity of CT45A1 in the advanced-stage (III-IV) cervical cancer patients reached 98% and 91%, respectively. These data indicate that CT45A1 is a new biomarker for the diagnosis of cervical cancer.
Fig. 1
CT45A1 is overexpressed in tumor tissues from cervical cancer patients and promotes tumor growth and metastasis in xenograft mice. Immunohistochemical staining showed that CT45A1 was overexpressed in the tumor tissues of cervical cancer patients (A-1 and A-2), but there was very little expression in the para-cancerous tissues (A-3 and A-4) and in the benign uterine myoma tissues (C). The results were scored and statistically analyzed (B). The CT45A1 expression levels in uterine myoma, cervical cancer stages I/II, and cervical cancer stages III/IV were also statistically analyzed (D). The correlation between CT45A1 levels and the overall survival of cervical cancer patients was assessed by gene expression profiling in the TCGA cohort and Kaplan–Meier analysis (E). The expression levels of CT45A1 in the organs and tissues of healthy individuals were measured by RT-PCR (F). Cervical cancer Caski cells transfected with either CT45A1-vector (CT45A1) or vector as a control (Vector) were subcutaneously injected into nude mice (n = 5/group). Tumor infiltration in each group was shown in G; the blue and pink circle points to the junction between the subcutaneous tumor and the peritoneum. The tumor volume was calculated (H). The tumors were weighted and analyzed (I). Data represent the mean (± SE) of the tumor vascular diameter in 17 tumor tissue fields of five tumor-bearing mice (J). The blood vessel number in the tumors with CT45A1 expression (K, tumor) was greater than in the tumors without CT45A1 expression; the red arrows refer to the blood vessels in the tumor. P values calculated by the log-rank test. Data are shown as the mean ± SE. *p < 0.05, **p < 0.01 in an unpaired t-test
Additionally, the t-test showed a significant positive correlation between CT45A1 levels and pathological cancer grade (Fig. 1D, p < 0.001, supplementary Tables S2 and S3). More importantly, Kaplan–Meier plots indicated that high expression of CT45A1 was associated with a poor prognosis in cervical cancer patients (kmplot.com) (Fig. 1E). The average survival time was shortened by 2.5 years in the cervical cancer patients with high CT45A1 levels as compared to those with low CT45A1 levels (p = 0.035). Furthermore, CT45A1 expression was investigated in 12 organs or tissues from healthy people by RT-PCR (Fig. 1F) and Real-time PCR (Supplementary Fig. S1B), respectively. The results showed that CT45A1 was overexpressed in the male testis but was not expressed or had extremely low expression in other normal tissues. Together, these data suggest that CT45A1 has high tumor specificity and sensitivity and is a new biomarker for the diagnosis and prognostic prediction of cervical cancer.
Next, the effect of CT45A1 on cervical tumor growth was examined in xenograft mice. Nude mice (n = 5/group) were subcutaneously injected with Caski cells with or without CT45A1 expression (Fig. 1G, Supplementary Fig. S2A, S2B). Forty-five days later, the tumor volume and weight were significantly increased in the CT45A1 expression group compared to the control group (Fig. 1H, I). Strikingly, the tumors with CT45A1 expression exhibited irregularly shaped edges and were invaded into the deep skin layer (Fig. 1G); the tumors in three out of five mice had disseminated to the peritoneum. By contrast, the tumors without CT45A1 expression had smooth surfaces or soft textures and were easily stripped from the skin layer. H&E staining indicated that the number of blood vessels was increased 3.5-fold in the tumors with CT45A1 expression compared to the control tumors without CT45A1 expression (Fig. 1J, K), suggesting that CT45A1 enhances tumor growth. Additionally, we performed in vitro tube formation assay to assess the effect of CT45A1 on tumor cell-mediated neovascularization. The result showed that the numbers of tube-like structures in CT45A1-overexpressing cervical cancer Caski cells were more than that of the control Caski cells without CT45A1 expression (Supplementary Fig. S1C and S1D), suggesting that CT45A1 promotes tumor cells-mediated neovascularization. In short, these data imply that CT45A1 enhances tumor malignant progression and neovascularization.
Moreover, CT45A1 significantly increased the migration and invasion of both cervical cancer Caski and Siha cells (Fig. 2A–H); convincingly, silencing of CT45A1 resulted in a significant decrease in HeLa cell migration and invasion (Fig. 2I–L), implying that CT45A1 increases cervical cancer cell motility. Additionally, a colony formation assay showed that CT45A1 increased the cervical cancer cell colony number 2.6-fold (Fig. 2M–P), suggesting that CT45A1 increases cervical cancer cell tumorigenesis. Collectively, these data indicate that CT45A1 enhances tumor growth, neovascularization, and metastasis in vivo and promotes cervical cancer cell tumorigenesis, tube-like structure formation, migration, and invasion in vitro.
Fig. 2
CT45A1 enhances cervical cancer cell migration, invasion and colony formation. CT45A1 promoted the migration of cervical cancer Caski (A, C) and Siha cells (E, G) and also enhanced the invasion of cervical cancer Caski (B, D) and Siha cells (F, H). By contrast, silencing of CT45A1 in HeLa cells inhibited cell migration (I, K) and invasion (J, L). The colony numbers in CT45A1-overexpressed Caski cells (M, CT45A1) and CT45A1-silenced HeLa cells (N, shRNA) were counted and statistically analyzed (O and P). Data are shown as the mean ± SE of three independent replicates. *p < 0.05, **p < 0.01 in an unpaired t-test
2.2 CT45A1 triggers the overexpression of oncogenic genes and activates tumorigenic signaling pathwaysDNA microarray showed that CT45A1 up-regulated the expression of 128 genes in cervical cancer Caski cells, including 68 pro-tumorigenic genes, such as fibronectin-1 (FN1), OXTR, PLAC8, LCP1, DACT1, KIAA1462, COL4A1, ABCA1, CNN1, GRB10, TNC, LMCD1, CPE, PLAC8, GNB4, TGFBI, LTBP1, CHML, KRT8, and COL4A2 (> twofold, p < 0.05, Fig. 3A, Supplementary Table S4). In particular, the expression level of tumorigenic FN1 was increased 14-fold in cervical cancer Caski cells with CT45A1 expression compared to control cells without CT45A1 expression (Fig. 3B). Many other oncogenic genes, including PLAC8, DACT1, KISS1, and GRB10, were also markedly increased (Supplementary Fig. S2C). In contrast, CT45A1 down-regulated 126 genes (> twofold, p < 0.05, Supplementary Table S4). Additionally, CT45A1 overexpression changed multiple signaling pathways in cervical cancer cells, including the ECM-receptor interaction, focal adhesion, Hippo, and PI3K-AKT signaling pathways (Supplementary Fig. S2D).
Fig. 3
CT45A1 upregulates fibronectin-1 (FN1) in cervical cancer cells. DNA microarray analysis showed CT45A1 induced differential expression of genes between Caski cells with and without expression of CT45A1; asterisk indicates that FN1 is the most up-regulated gene among the CT45A1-regulated genes (A). QT-PCR confirmed that CT45A1 markedly increased FN1 mRNA levels (B). Western blot showed that the overexpression of CT45A1 notably increased FN1 protein levels in Caski cells and in the supernatant of the cell culture (SPN FN1) (C), and the data were statistically analyzed (D and E). The FN1 gene promoter region (-1354 bp to + 247 bp) and the location of the transcription factor CREB are shown (F). The luciferase assay indicated that CT45A1 markedly increased FN1 promoter activity (G). Silencing of CT45A1 reduced FN1 protein levels (H, I) and decreased Caski-CT45A1 cell migration (J, K). Computer analysis predicates potential CT45A1 protein-binding site 1 and site 2 in the FN1 gene promoter region (L). ChIP showed that CT45A1 bound to the site 1, but did not bind to the site 2 in FN1 promoter region (M). QT-PCR also indicated that CT45A1 bound to the site 1 (N). The site 1 and site 2 nucleic acids, and control nucleic acids were spotted on nitrocellulose membranes, and blocked with 5% Nonfat-Dried Milk buffer. After incubation with CT45A1 protein, the binding of CT45A1 protein to FN1 gene promoter site 1 and site 2 nucleic acids, and control nucleic acids were detected by CT45A1 specific monoclonal antibody and Dot blot (O). Data are shown as the mean ± SE of at least three independent replicates. *p < 0.05, **p < 0.01 in an unpaired t-test
Further investigation revealed that CT45A1 markedly elevated both FN1 mRNA and protein levels in cervical cancer cells (Fig. 3B–E, Supplementary Fig. S3A and S3B). Mechanistically, CT45A1 promoted FN1 gene transcription by enhancing FN1 gene promoter activity (Fig. 3F and G) and interacting with the transcription factor CREB (Supplementary Fig. S3C-S3I). By contrast, the Caski cells that overexpressed CT45A1 was silenced by shRNA significantly reduced FN1 levels (Fig. 3H and I) and inhibited Caski cell migration (Fig. 3J–K). Together, these data suggest that CT45A1 regulates FN1 gene transcription in cervical cancer cells.
CT45A1 is a nuclear protein. We recently identified the CT45A1 protein-binding consensus sequence CGCCCC (Fig. 3L). In the current study, we first explored whether the CT45A1 protein-binding CGCCCC exists in the FN1 gene promoter region (Fig. 3L). The result showed that there are two CGCCCC sequences in the FN1 gene promoter region (Fig. 3L), named as the site 1 and site 2, respectively. Next, the direct binding between purified CT45A1 recombinant protein and these two sites GATCCGAAAGCGCCCCGCGGAATCT (site 1) and TCTCTCCCCCCCGCCCCG GGCCTCCAG (site 2) was accessed by CHIP. The results showed that CT45A1 directly bound to the site 1 in cervical cancer cells but did not bind to the site 2 (Fig. 3M–N, Supplementary Fig. S2E). Convincingly, Protein-DNA binding Dot blot confirmed the results (Fig. 3O), suggesting that although the core nucleic acid sequence CGCCCC is important for CT45A1 binding, the front and downstream nucleic acids of the CGCCCC is also critical for the binding of CT45A1 protein to the FN1 gene promoter. In brief, these data indicate that CT45A1 binds to the FN1 gene promoter and drives transcription of the gene.
We next examined whether CT45A1 affects the FN1 downstream oncogene SRC. Western blotting showed that CT45A1 overexpression significantly increased SRC phosphorylation, whereas silencing of CT45A1 in Caski-CT45A1 cells reduced SRC phosphorylation (Fig. 4A, B). Notably, silencing of FN1 by shRNA completely abolished CT45A1-induced SRC activation (Fig. 4C and D), implying that FN1 is at the downstream of CT45A1.
Fig. 4
CT45A1 activates the oncogene SRC via interaction with the protein in cervical cancer cells. Caski cells barely expresses CT45A1. We first transfected Caski cells with CT45A1 cDNA-vector and empty vector, respectively, to produce CT45A1 overexpressed Caski cells, then CT45A1-overexpressed Caski cells were silenced by shRNA. Western blot indicated that the overexpression of CT45A1 triggered SRC phosphorylation in cervical cancer cells (A, B). Silencing of fibronectin 1 (FN1) notably reduced SRC phosphorylation (C, D). Co-immunoprecipitation showed that CT45A1 interacted with SRC (E, F). Immunofluorescence staining revealed co-localization of SRC with CT45A1 in the nucleus; red refers to SRC, green refers to CT45A1, and light blue arrows refer to changes in the SRC location (G and H, 600 ×). Pull down of HeLa cell lysate with CT45A1-specific antibody further confirmed the interaction between CT45A1 and SRC (I). In vitro protein kinase assay showed that CT45A1 induced SRC phosphorylation in an ATP-dependent manner (J, K). Data are shown as the mean ± SE of three independent replicates. *p < 0.05, **p < 0.01 in an unpaired t-test
Additionally, co-immunoprecipitation (Co-IP) revealed that CT45A1 was able to directly bind to the SRC protein in cervical cancer cells (Fig. 4E, F). Immunofluorescence imaging indicated that CT45A1 changed the SRC protein localization from the sub-cellular membrane to the cytoplasm and nucleus in cervical cancer cells (Fig. 4G, H). Furthermore, the direct binding between CT45A1 and SRC proteins was further confirmed by a pulldown assay (Fig. 4I). Moreover, an in vitro protein kinase activity assay showed that CT45A1 markedly increased SRC protein kinase activity in an adenosine triphosphate (ATP)-dependent manner (Fig. 4J, K). Convincingly, the SRC-specific inhibitor 6-dimethylamino-2-phenyl-3(2H)-pyridazinone (PP2) markedly suppressed CT45A1-mediated SRC-ERK-CREB activation (Fig. 5A–D) and inhibited cervical cancer cell migration (Fig. 5E and F). Additionally, silencing of CT45A1 in HeLa cells significantly reduced the levels of oncogenic FN1, p-SRC, p-ERK, and p-CREB (Supplementary Fig. S4A-4E). Together, there data suggest that CT45A1 is a new activator of oncogenic SRC, and there is a novel pro-tumorigenic CT45A1-FN1-SRC-ERK-CREB signaling pathway in cervical cancer (Fig. 5G), importantly, CT45A1 is at the front of the signaling pathway; and silencing of CT45A1 inhibits multiple oncogenic signaling pathways.
Fig. 5
The CT45A1-SRC-ERK-CREB axis controls the migration of cervical cancer cells. The SRC inhibitor PP2 (5 μM) and ERK inhibitor SCH772984 (5 μM) reduced the phosphorylation of SRC, ERK, and CREB (A-D). CT45A1-induced cervical cancer cell migration was significantly inhibited by the SRC inhibitor PP2 (E, F). Schemes of the mechanism underlying the regulation of the migration by the newly identified CT45A1-SRC-ERK-CREB axis (G). Data are shown as the mean ± SE of three independent replicates. *p < 0.05, **p < 0.01 in an unpaired t-test
Next, the effects of CT45A1 on the expression of the oncogenic proteins Yes-associated protein (YAP)/ tafazzin (TAZ) downstream of the Hippo signaling pathway were investigated. Western blotting revealed that the levels of the YAP/TAZ proteins were notably increased in cervical cancer Siha cells with CT45A1 expression (Fig. 6A–C), but did not significantly affect several other signaling pathways (Supplementary Fig. S4F and S4G). By contrast, silencing of CT45A1 in HeLa cells by shRNA markedly reduced the levels of the YAP/TAZ proteins (Fig. 6D–F), whereas the YAP and TAZ mRNA levels were not significantly changed (Supplementary Fig. S4H). Additionally, CT45A1 interacted with the YAP and TAZ proteins (Fig. 6I–K) and co-localized with YAP and TAZ in the nucleus (Fig. 6L, Supplementary Fig. S4I-S4K). Convincingly, the SRC inhibitor PP2 abolished CT45A1-induced YAP expression (Fig. 6G and H). These data suggest that CT45A1 is a new inducer of tumorigenic YAP/TAZ, and there is a new oncogenic CT45A1-SRC-YAP/TAZ signaling pathway in cervical cancer cells. Collectively, CT45A1 plays an important role in triggering tumorigenesis and is a target for anti-cervical cancer therapy.
Fig. 6
CT45A1 activates the oncogenic proteins YAP and TAZ in cervical cancer cells. Western blot revealed that CT45A1 overexpression increased the levels of the oncogenic YAP and Tafazzin (TAZ) proteins in Siha cells (A-C). By contrast, silencing of CT45A1 in HeLa cells diminished the expression of YAP and TAZ (D-F). Additionally, the SRC inhibitor PP2 significantly suppressed CT45A1-induced expression of YAP (G, H). Co-IP (I, J) and pull-down assays (K) revealed that CT45A1 interacted with YAP and TAZ. Immunofluorescence staining and confocal microscopy techniques confirmed that YAP and CT45A1 were co-localized in the nucleus; red refers to YAP and green refers to CT45A1 (L, 1200 ×). Data are shown as the mean ± SE of three independent replicates. *p < 0.05, **p < 0.01 in an unpaired t-test
2.3 CT45A1 boosts cisplatin drug resistance and apoptosis resistance and is a target for developing novel therapeutics against cervical cancerOverexpression of CT45A1 was found to boost cisplatin resistance and apoptosis resistance (Fig. 7) in cervical cancer Siha cells. CT45A1 significantly diminished 10 μM cisplatin-induced DNA damage (Fig. 7A–E), as evident by decreases in levels of the DNA damage marker γH2AX (Fig. 7B, C and H) and apoptotic cells (Fig. 7F and G). This suggests that CT45A1 increases cervical cancer cell cisplatin drug resistance and apoptosis resistance.
Fig. 7
CT45A1 enhances cisplatin drug resistance and apoptosis resistance in cervical cancer cells. CT45A1-expressing Siha cells were treated with 0–20 μM of the anti-cancer drug cisplatin for 48 h and cell proliferation was assessed with Alarm blue assays (A). The levels of cleaved PARP, γH2AX, and caspase 3 (cas3) in CT45A1-expressing Siha cells were compared to the control Siha cells without expression of CT45A1 (C-E). The apoptotic assay indicated that apoptosis in CT45A1-expressing Siha cells was significantly reduced when the cells were treated with cisplatin for 48 h (F, G). Immunofluorescence staining and confocal microscopy techniques confirmed that the expression of γH2AX was diminished in CT45A1-expressing cells as compared to control cells without expression of CT45A1; red refers to γH2AX and blue refers to DAPI (H, 2500 ×). Data are shown as the mean ± SE of three independent replicates. * p < 0.05, ** p < 0.01, and *** p < 0.001 in an unpaired t-test
Next, CT45A1-targeted therapeutics were explored and the small molecule lycorine (MW: 287.31) was found to markedly reduce CT45A1 levels in cervical cancer cells (Fig. 8A, B). Additionally, lycorine inhibited the phosphorylation of oncogenic SRC and ERK in a concentration-dependent manner (Fig. 8A, C, D), reduced cervical cancer cell colony numbers (Fig. 8E, F), and decreased HeLa cell invasion (Fig. 8G, H). Furthermore, we investigated the effect of lycorine on expression of oncogenic YAP and TAZ in cervical cancer HeLa cells. The results showed that after treatment of HeLa cells with lycorine for 72 h, YAP levels in the cells were significantly reduced by lycorine at concentrations of 20 and 40 μM; meanwhile TAZ levels were also significantly diminished by lycorine at the concentration of 40 μM compared to the control without lycorine treatment (Supplementary Fig. S5A-S5C). These data indicate that lycorine reduces CT45A1-induced overexpression of oncogenic YAP in cervical cancer cells, implying that lycorine is a new inhibitor of the CT45A1-SRC-YAP signaling pathway.
Fig. 8
Reduction of CT45A1 expression by lycorine suppresses tumor growth of cervical cancer cells. CT45A1-overexpressing HeLa cells were treated with the small molecule lycorine (LH) at concentrations of 0–40 μM for 72 h. The protein levels of CT45A1, phosphorylated-SRC (p-SRC), and phosphorylated-ERK (p-ERK) were measured by western blot (A) and were statistically analyzed (B-D). HeLa cell-forming colonies were counted (E, F). Lycorine significantly inhibited HeLa cell invasion (G, H) and tumor growth in tumor-bearing nude mice (I, J, n = 6 in each group). H&E staining showed that the number of tumor blood vessels was significantly reduced (K); the red arrows refer to the blood vessels, and the data represent the tumor vascular diameter mean (± SE) of six mice (L). Additionally, CT45A1-overexpressing HeLa cells were first treated with lycorine at the concentration of 5 μM for 72 h, the living cells were counted and added to the 48-well plates coated with Matrigel. The tube-like structures in the randomized fields were imaged (M), counted and statistically analyzed (N). Data are shown as the mean ± SE of three independent replicates. *p < 0.05, **p < 0.01 in an unpaired t-test
In the xenograft mouse model, lycorine treatment of tumor-bearing mice significantly reduced the tumor volume (Fig. 8I) and tumor weight (Fig. 8J) and decreased the number of blood vessels 2.3-fold (Fig. 8K and L) as compared with the saline control. Strikingly, tube formation assay showed that the tube forming ability of CT45A1-overexpressing HeLa cells was completely inhibited by 5 μM lycorine (Fig. 8M and N). There were no obvious complications in the mice treated with lycorine at the effective dosage (Supplementary Fig. S5D and S5E). Together, these data indicate that CT45A1-enhanced tumorigenesis, neovascularization, cisplatin drug resistance, and apoptosis resistance can be effectively reduced by lycorine, and lycorine is a new CT45A1 expression suppressor and a novel cervical cancer inhibitor. Conceptually, these findings indicate that inhibition of CT45A1 expression is a new strategy for cervical cancer therapy.
留言 (0)