
Remember me

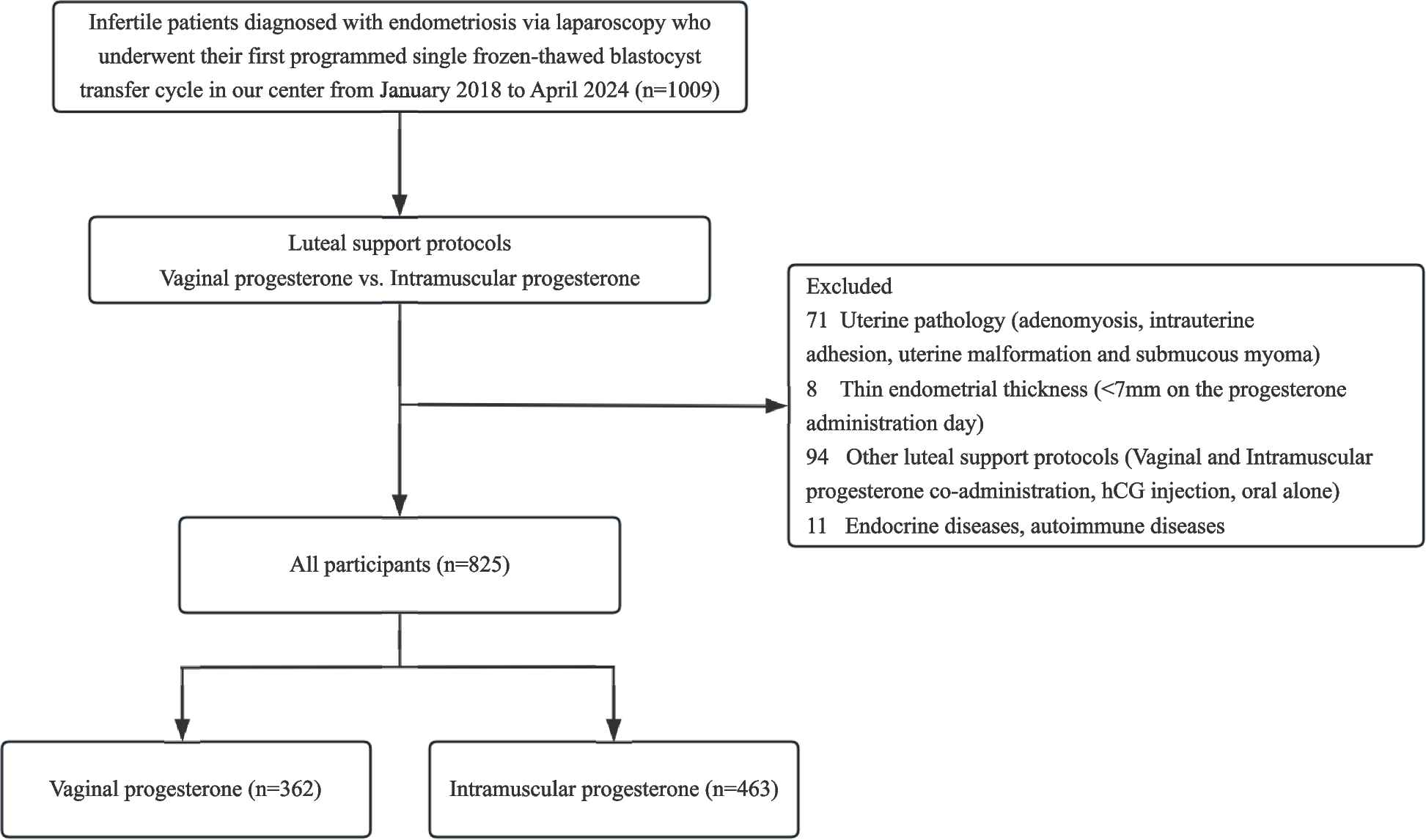







The biogenesis of miRNA initiates from the nucleus where miRNA genes get transcribed into long hairpin primary transcripts. Drosha along with its cofactor DGCR8 forms a complex called microprocessor and processes primary transcripts into small hairpin RNAs (pre-miRNA) (Fig. 1). Following Drosha processing, pre-miRNA is transported to the cytoplasm for further processing through a protein, Exportin-5. In the cytoplasm, pre-miRNA is cleaved by another protein called Dicer to yield a small RNA duplex. This small RNA duplex is loaded onto an AGO protein, forming a complex called RNA-induced silencing complex (RISC), which unwinds the RNA. The unwinding of the duplex generates two strands, passenger strand (miRNA*) and guide strand. miRNAs can target multiple mRNA molecules in a specific manner, which makes miRNAs powerful regulatory molecules [35].
Fig. 1
Biogenesis of small non-coding RNAs
miRNAs in PGCsIn the primordial germ cells (PGCs), miRNAs regulate a number of pluripotency genes involved in germ cells specification. In human embryonic stem (ES) cells, miR-145 completely suppresses the expression of OCT4 and partially suppresses SOX2 expression and hence promotes differentiation. The deletion of Drosha and Dicer in male PGCs disrupted its propagation, differentiation, and maturation [36]. PGCs start to differentiate from embryonic stages E11.5 stage and at E13.5 female PGCs enter meiosis while male PGCs enter mitotic arrest in mice. It has been reported that several miRNAs play an important role in gonad sexual determination. It was reported that expression of miR-103-3p, let-7 g-5p, miR-107-3p, and miR-26a-5p was down-regulated in E11.5 male somatic cells [37]. These miRNAs target key genes (Cyp26b1, Fgf9) in male germ cell differentiation. Moreover, eighteen miRNAs (mmu-miR-19a-3p, mmu-miR-22-3p, mmu-miR-30a-5p, mmu-miR-30d-5p, mmu-miR-30e-5p, mmu-miR-125a-3p, mmu-miR-139-5p, mmu-miR-140-5p, mmu-miR-149-5p, mmu-miR-185-5p, mmu-miR-202-3p, mmu-miR-204-5p, mmu-miR-214-3p, mmu-miR-500-3p, mmu-miR-532-3p, mmu-miR-532-5p, mmu-miR-665-3p, mmu-miR-667-3p) were up-regulated in E13.5 male somatic cells, which target genes related to activin signaling pathway, and activin is known to inhibit Cyp26B1 [37]. They also found two clusters of miRNAs (miR-199-214, miR-182-183-96) to be differentially expressed in embryonic stages E11.5, E12.5 and E13.5, whose targets are involved in gonad development [37]. Male germline committed PGCs showed up-regulation of let-7 family, miR-125a, and miR-9 [36], whereas miR-29b was found to be up-regulated in female germ line committed PGCs, where it epigenetically regulates gene expression by targeting Dnmt3a and Dnmt3b [38].
miRNAs in spermatogenesisSpermatogenesis is the main and crucial step of male reproduction, which requires an accurately, spatially and temporally regulated miRNA expression pattern [39]. To maintain the germ cells population throughout sexual maturity, the spermatogonial stem cells (SSCs) must divide in order to self-renew the stem cell pool. Through high-throughput sequencing, it was revealed that a number of miRNAs contribute to the regulation of SSC status. Recently, many studies investigating the role of small RNAs in spermatogenesis have revealed the importance of miRNAs during this process. Zhou et al. [40] reported that miR-663a is important for the proliferation of SSCs [40]. They found NFIX to be the direct target of miR-663a and that silencing of miR-663a leads to an increase in the proliferation rate and late apoptosis of human SSCs in-vitro [40]. In another study, miRNA functional assays suggested that miR-322 is critical for SSC self-renewal by targeting RASSF8 (ras association domain family 8) and miR-100 promotes the proliferation of spermatogonial stem cells via regulating Stat3 [41, 42]. The fate of SSCs is mainly determined by genetic and epigenetic factors. Similarly, molecular studies on the SSCs revealed that miRNA-31-5p regulates the proliferation, DNA synthesis, and apoptosis of human SSCs in association with PAK1-JAZF1-cyclin A2 pathway [43]. Liu et al. [44] demonstrated that Chd1 (chromo domain helicase/ATPase DNA binding protein 1-like) mediated downregulation of miR-486 promotes self-renewal potential of SSCs via MMP2 (matrix metalloproteinase 2) regulation [44].
Tan et al. [11] undertook next-generation sequencing to compare small RNA signatures between different cell populations like SSC, Sertoli cells, developing germ cells, and mouse embryonic stem cells grown in vitro [11]. It was concluded that miRNA and endo-siRNA expression levels decline as spermatogenesis proceeds while piRNA expression elevates. Apart from finding several novel miRNAs expressed in mouse SSCs, the group also reported miRNAs common between the SSCs and embryonic stem cells (ESCs) in order to understand the molecular mechanism involved in the conversion of SSCs to ESCs known as germline-derived pluripotent stem cells [11]. In another sequencing study, miRNA expression profiling was conducted in Thy1+ cell population, which is highly enriched in SSCs, and Thy1− cell population, composed mainly of testicular somatic cells. After comparing the two groups, they found that miR-21, miR-34c, miR-182, miR-183, and miR-146a were preferentially expressed in the Thy1+ cell population compared with Thy1− somatic cells [45]. They further demonstrated that the inhibition of miR-21 in SSC-enriched germ cell culture increased the rate of apoptosis in germ cells; therefore, it is critical for the survival of SSC population [45]. Despite these advancements, the functions and mechanisms of a number of miRNAs in regulating stem cell renewal and differentiation are yet to be explored. He et al. [46] reported that miRNA-20 and miRNA-106a were preferentially expressed in mouse SSCs. In-silico miRNA target prediction reported, STAT3 and CCND1 as the direct targets of miRNA-20 and miRNA-106a. Furthermore, the knockdown of Stat3 and Ccnd1 lead to SSC renewal [46]. Similarly, in another study, it was reported that miR-202 is the key regulatory miRNA for SSCs renewal and maintenance. The function of miR-202 was exploited by knockdown through CRISPR-Cas9 technology and it was found that the absence of miR-202 could lead to early differentiation, reduced stem cell property, and increased mitosis and apoptosis [47]. Through iTRAQ-based proteomic analysis and RNA sequencing, the target genes were identified, which were mainly cell cycle regulators and RNA binding proteins. It was further reported that miR-202 targets Rbfox2 and cpeb1, which are responsible for the differentiation of SSCs into meiotic cells [47].
miR-122a, miR-18a, and miR-34c are some of the crucial miRNAs regulating pivotal steps in spermatogenesis [48]. In the male germ cells, miR-122a is expressed in the late-stage germ cells and it down-regulates the expression of transition protein 2 (Tnp2) post-transcriptionally, a protein involved in replacing histones during chromatin condensation [48]. miR-18a belongs to the miR-17-92 cluster and is associated with the regulation of genes involved in cancer [49]. miR-18a is an important miRNA for spermatogenesis as it directly targets heat shock factor 2 (Hsf2), a transcription factor required for the regulation of many genes involved in spermatogenesis and embryogenesis [48, 50]. miR-17-92 cluster consists of miRNAs from four different families (miR-17, miR-18, miR-19, and miR-25). Knockout studies on mir-17-92 cluster have shown reduced testis weight and size and some seminiferous tubules with only Sertoli cells, but the mice were fertile [51]. The above-mentioned targeted disruption of miR-17-92 in the testes of adult mice gives rise to severe testicular degeneration, empty tubules and low sperm production. This phenotype is due to the reduced number of spermatogonia and spermatogonial stem cells (SSCs), and significantly increased germ cell apoptosis in the testes of miR-17-92-deficient mice. All the above suggest that miR-17-92 is essential for normal spermatogenesis in mice [52]. Recently, a study has shown a significantly reduced fertility in miR-17 ~ 92+/− and miR-106b-25−/− double mutant male mice due to oligozoospermia and disrupted spermatogenesis [53]. In another study, miR-469 was found to be up-regulated in GRTH/DDX25 null mice where it was reported to target Tnp2 and Prm2, which ultimately resulted in infertility due to failure in the generation of elongated spermatids [54]. Recently, X-linked miRNAs were evaluated in highly purified spermatogenic cells at different stages, finding that X-linked spermatogenesis-related miRNAs (SpermiRs) expressed in early meiotic phase and showed a conserve testis specific expression in mammals [55]. This study revealed the compensatory upregulation of miR-465a-5p, after knockdown of another abundant miRNA (miR-741) in cultured mouse SSCs, which did not affect the genome-wide mRNA expression levels in cell line. Ota et al. [56] found that X-linked miR-871 and miR-880 cooperatively regulate spermatogenesis via the WNT/β-catenin pathway during testicular germ cell development by targeting Fzd4 (WNT receptor), which further indicates that WNT/β-catenin signaling promotes proliferation of SSCs, but represses their differentiation. Double mutant mice having deletion covering miR-871 and miR-880 resulted in meiotic arrest in a few seminiferous tubules [56].
In a recent study, miR-202-5p was found to be critical to spermatogenesis as its in-vivo inhibition showed spermatogenic arrest in murine testis seminiferous tubules [57]. MiR-34 family comprises of six members (miR-34a, miR-34b, miR-34c, miR-449a, miR-449b, miR-449c) located on three different chromosomes 1p36.22, 11q23.1, and 5q11.2. It has been reported that miR-34 family has significant role in spermatogenesis owing to its high expression in the male germ cells [58, 59]. The members of this family are expressed in spermatogonial stem cells (SSCs) and as well as spermatozoa. The expressions of these family members shoot up in the beginning of meiosis, suggesting their role in spermatocytes [60]. The miR-34 family mainly targets the genes of cell cycle, such as Notch1, Cdk4, and Myc [61]. Interestingly, disruption of a few members of this family led to an increase in the expression of other members showing compensatory effects [58]. Wu et al. [59] found that miR-34b/c and miR-449 clusters are functionally redundant. They inactivated both the clusters simultaneously and found disruption of their target genes involved in spermatogenesis, brain development, and microtubule dynamics. MiR-34c is also present in the pachytene spermatocytes and round spermatids [59]. Tgif2 and Notch are the two transcription factors important for spermatogenesis that are direct targets of miR-34c [61]. MiR-34c along with other miRNAs, such as miR-449 and 34b lead to apoptosis of germ cells by targeting BCL2 and ATF1 [62, 63].
miRNAs in male infertilitySince miRNAs play critical roles in gene regulation during spermatogenesis, their alterations could lead to male infertility. It is interesting to note that semen contains extracellular and intracellular small RNAs. miRNAs have been thoroughly investigating to have a significant link to numerous facets of male infertility (Table 1). The intracellular small RNAs might have originated in the testis during spermatogenesis or during their maturation in the epididymis. Spermatozoa are well known to lose and gain small RNAs during their transit through epididymis. Apart from intracellular small RNAs, seminal fluid has cell free small RNAs, which might have originated in the testis, epididymis or in male accessory glands. Both the intracellular and cell free small RNAs have been investigated for their roles in fertility and their correlation with infertility. In the following sections, we will discuss testicular miRNAs, sperm-borne miRNA, epididymal small RNAs to highlight their importance in spermatogenesis, fertility and infertility (Fig. 2).
Table 1 Studies on differential expression of miRNAs in male infertility Fig. 2
miRNAs critical to various stages of germ cell development in rodents. ?- represents the lack of study on testicular sperm miRNAs
Testicular miRNA in male infertilityTestes consist of an impressive signature of miRNAs, which regulate several important biological pathways critical for the process of spermatogenesis. In the very first study, Barad et al. 2004 used microarray to identify miRNA signatures in five different tissues, i.e., liver, brain, thymus, placenta, and testis, and found abundant expressions of five miRNAs (let-7a, miR-10b, miR-134, miR-187 and miR-212) in the human testicular tissue [64]. After a few years, a microarray study was conducted by Lian et al. [65] on testicular biopsies of three non-obstructive azoospermic patients and two testicular biopsies were taken from the patients undergoing orchiectomy for prostrate carcinoma. They identified 154 down-regulated and 19 upregulated miRNAs in non-obstructive azoospermic (NOA) patients as compared to controls [65]. Abu-Halima et al. [66] conducted yet another microarray on testicular tissues of infertile men with different histopathologic patterns. Forty infertile patients were recruited, of which 12 were Sertoli cell only syndrome (SCOS), 12 had mixed atrophy (MA) and 16 had germ cell arrest at spermatocyte stage (GA) while sixteen individuals with normal spermatogenesis and normal structure of seminiferous tubules were recruited as controls. They found a total of 197, 68, and 46 differentially expressed miRNAs in SCO, MA and GA patients, respectively as compared to controls [66]. miR-449 family (miR-449a and miR-449b*) and miR-34 family (miR-34b*, miR-34b and miR-34c-5p) were found to be highly down-regulated in SCO, MA, GA groups as compared to controls. In normal spermatogenesis, these miRNAs are abundantly expressed in testis, suggesting that these two miRNA families are crucial for spermatogenesis [66]. Yang et al. [67] identified 770 miRNAs in normal testis tissue using NGS based technology for the first time. Another group of scientists reported the association of miR-302 cluster, miR-21, miR-29a and miR-367 with testicular germ cell tumor cases [60].
Noveski et al. [68] conducted microarray analysis to identify miRNA expression in 27 hypospermatogenesis (HS) men, three hypospermatogenesis men with AZFc deletions, eight SCO, two MA, eight testicular atrophy/fibrosis/hyalinization (TFH) and eight 46,XXY men as compared to 18 obstructive azoospermic men recruited as controls. They identified six miRNAs to be down-regulated in patients with AZFc deletions, 27 miRNAs to be up-regulated and 3 down-regulated in MA patients, 32 miRNAs to be up-regulated and 20 miRNAs to be down-regulated in SCO patients as compared to controls. Five of the promising miRNAs (miR-34b, miR-449b, miR-517c, miR-181c, and miR-605) were validated by qRT-PCR. In another microarray study, Zhang et al. [69] identified 129 miRNAs to be differentially expressed in testicular tissues of NOA patients as compared to controls. Eight miRNAs were selected for validation by qRT-PCR, of which four were up-regulated (miR-370-3p, miR-10b-3p, miR-539-5p, miR-22-5p) and four were down-regulated (miR-34b-5p, miR-31-5p, miR-516b-5p and miR-122-5p) [69]. Apart from elucidating the miRNA expression in infertile testicular tissues, Yao et al. [12] conducted next generation sequencing to identify miRNA expression in isolated spermatogonia, pachytene spermatocytes and round spermatids from 60 NOA patients and 20 controls using STA-PUT. On further comparing the miRNA expression profile, they found a total of 396 DE miRNAs between spermatogonia of OA and NOA groups (88 miRNAs up-regulated and 308 miRNAs down-regulated), 395 DE miRNAs between pachytene spermatocytes of OA and NOA groups (97 miRNAs up-regulated and 298 miRNAs down-regulated) and 378 DE miRNAs between round spermatids of OA and NOA groups (64 miRNAs up-regulated and 314 miRNAs down-regulated) [12]. A recent study by Piryaei et al. [70] showed downregulation of 120 miRNAs and upregulation of 10 miRNAs in NOA cases when compared to OA. The top 10 downregulated miRNAs included hsa-miR-449a, hsa-miR-34c-3p, hsa-miR-375, hsa-miR-517b-3p, hsa-miR-512-3p, hsa-miR-34c-5p, hsa-miR-520c-3p, hsa-miR-516b-5p, hsa-miR-1323, hsa-miR-34b-3p [70]. Research done so far has concluded a significant link between altered expression of miR-34/449, let-7, miR-517 and spermatogenic disruption, leading to infertility in male (Table 1) (Fig. 3).
Fig. 3
Differentially expressed small non-coding RNAs in primordial germ cells (PGCs), testis, seminal plasma, and sperm of infertile men. Parts of the figure were drawn using pictures from Servier Medical Art. Servier Medical Art by Servier is licensed under a Creative Commons Attribution 3.0 Unported License (https://creativecommons.org/licenses/by/3.0/ accessed on 15 January 2023)
Epididymal miRNAsEpididymis has three distinct regions: caput, corpus and cauda. During sperm maturation, spermatozoa pass through the epididymis to acquire motility and fertilizing capability. In epididymis, sperm come in contact with different repertoires such as proteins and transcripts, which are released from the epithelium of epididymis via epididymosomes. It has been reported that sperm miRNA signatures change as they travel along the epididymis. Belleannѐe et al. [71] performed microarray in human epididymis from three donors to identify miRNA expression pattern in the epididymis of healthy males. They found 281, 282 and 289 miRNAs to be expressed in the caput, corpus and cauda region of epididymis, respectively, with 35 miRNAs showing differential expression across these segments of epididymis. Further, they found that five members of the miR-888 cluster (miR-890, miR-891a, miR-891b, miR-892a, and miR-892b) were abundantly expressed in caput and the corpus region, suggesting their role in the latter stages of sperm maturation [71]. In order to understand the role of miRNAs in epididymis during post-natal development, Zhang et al. [72] performed miRNA profiling in the epididymis of a new-born, 25 years old, and 75 years old males and found that epididymis of new born expressed more number of miRNAs whereas epididymis of 25 and 75 years old males expressed a few miRNAs, with 127 miRNAs exclusively expressed in the new-born epididymis whereas only 3 and 2 were exclusively expressed in the epididymis of 25 and 75 years old males, respectively. There were 106 miRNAs which were ubiquitously expressed in all the three ages, including let-7a, b, c, d, f, miR-125a, miR-125b, miR-143, miR-23b, miR-26a and rno-miR-347 [72]. In a case-control study, miRNA expression in epididymis of three control and three vasectomized patients was identified using microarray [71]. They identified a total of 336 and 394 miRNAs in the epididymis of vasectomized and control donors, respectively. Further, they identified four (miR-126, miR-129, miR-326 and miR-16), two (miR-1826 and miR-18a) and nine miRNAs (miR-941, miR-548b, miR-1826, miR-1281, miR-1825, miR-548c, miR-1278, miR-421 and miR-135a) that differed in expressions in caput, cauda and corpus, respectively after vasectomy. Additionally, they found a total of 22 miRNAs were down-regulated and 8 miRNAs were up-regulated in the epididymis of vasectomized patients as compared to controls [71]. Studies done so far have shown loading of 115 miRNA RNAs on spermatozoa and shedding of about 113 small RNAs from spermatozoa during the epididymal maturation process [73].
Recent investigations on the effect of stress on male fertility and transgenerational inheritance have shown a critical role of epididymis in altering the small RNA profile of spermatozoa, which may lead to a change in transcriptome of the zygote [74]. miR-31-5p was reported to be majorly altered miRNA along with miR-155-5p, miR-878-5p, and miR-34c-5p in cauda epididymosomes of adult males exposed to postnatal stress [74]. A number of other sperm-borne small RNAs have been shown to alter upon exposure to high fat diet, glucose intolerance, stress, resilience, and other endocrine disruptors [75]. These studies have provided evidences that these epididymis miRNAs may mediate transgenerational changes aimed at betterment of species.
Seminal plasma miRNAsSeminal plasma is constituted by the secretions of the testis, seminal vesicles, and prostate. A number of small RNAs leach out from the testicular cells or accessory glands to make their way into the seminal plasma. MicroRNAs are abundant and stable in seminal plasma, which can be exploited as biomarkers of fertility.
Seminal plasma has a large number of cell free small RNAs, which may originate in the testis, epididymis or male accessory glands. These small RNAs are often in extracellular vesicles, which have unknown significance. Till date, over 21,000 numbers of small RNAs have been reported in seminal plasma. While their biological significance remains hitherto unknown, their utilization as the markers of fertility and infertility has been tried by a number of studies. Extracellular small RNAs have particularly been interrogated in azoospermia cases where there are no sperm in the ejaculate.
Wu et al. [8] performed qRT-PCR on the seminal plasma of 48 NOA patients and 48 fertile controls and identified miR-19b and let-7a to be differentially up-regulated in NOA patients compared to fertile controls [8]. The same research group later performed TaqMan low-density arrays (TLDA) analysis to identify differential miRNA expression in 20 NOA patients and 20 fertile controls [76]. Two independent validations led to the identification of three miRNAs (miR-141, miR-429 and let-7-1-3p) that were significantly upregulated in NOA patients as compared to fertile controls. Further, the ROC curve analysis showed that these miRNAs could be used as semen-based biomarkers for diagnosis of NOA [76].
A few studies have identified altered miRNA expressions in the extracellular micro-vesicles present in the seminal plasma (Table 1). Barcelo et al. [77] undertook miRNA profiling analysis using qRT-PCR panels in 14 infertile azoospermic patients with spermatogenic failure and 13 obstructive azoospermic individuals with conserved spermatogenesis. They found that miR-31-5p could be useful in distinguishing between OA patients with conserved spermatogenesis from NOA patients with failed spermatogenesis. Additionally, they showed that the combined expression values of miR-539-3p and miR-941 could be useful in determining the presence of sperm in patients with severe spermatogenic disorders [
Comments (0)