Thalassemia is a prevalent genetic disorder worldwide that results from abnormal production of α/β globin due to mutations during mRNA translation [1]. The unpaired globin is seen in erythrocytes as cytoplasmic inclusion bodies and causes the shortening the erythrocytes' lifespan and occurrence of chronic hemolytic anemia. In intermediate β-thal and some hemoglobinopathies, elevation of HbF levels in adult erythroblasts is an important therapeutic target [1, 13, 14]. The mechanism by which HU generates her HbF is unknown. The discovery of associations between genes affected by HU and those involved in HbF switching may help explain some of these mechanisms.
In this study, using transcriptome analysis by WGCNA method, six genes were selected and the expression of these genes (IGF2Bp3, LIN28A, COL4A5, BCL2, and SP1) under HU treatment and their relationship with γ-globin switching were evaluated.
According to the findings, the level of expression for the LIN28A gene was elevated following HU treatment. The discovery of this gene initially occurred in the Caenorhabditis elegans nematode, and it is responsible for regulating the timing and progression of developmental processes [15]. In humans, two similar versions of the LIN28 gene, known as LIN28A and LIN28B, have been identified. These genes are involved in promoting pluripotency in stem cells and the differentiation of skeletal muscle cells. They also contribute to various physical developments such as changes in height, puberty timing, and age at menopause [15]. The LIN28 gene alters the let-7 family of miRNAs, and its expression plays a role in blocking let-7 [15]. In mature red blood cells, LIN28 protein has been found to reduce let-7 miRNA levels and potentially increase HbF expression [16]. Lee et al. confirmed that a higher level of LIN28B expression in mature erythroid cells can lead to down-regulation of let-7 expression and a significant increase in HbF expression level. Furthermore, they found that increased expression of the HbF in mature CD34 + cells coincided with a decrease of mature globin expression [17]. These findings align with Lee et al.’s results that LIN28A, homologous to LIN28B, also increases its expression under the influence of HU.
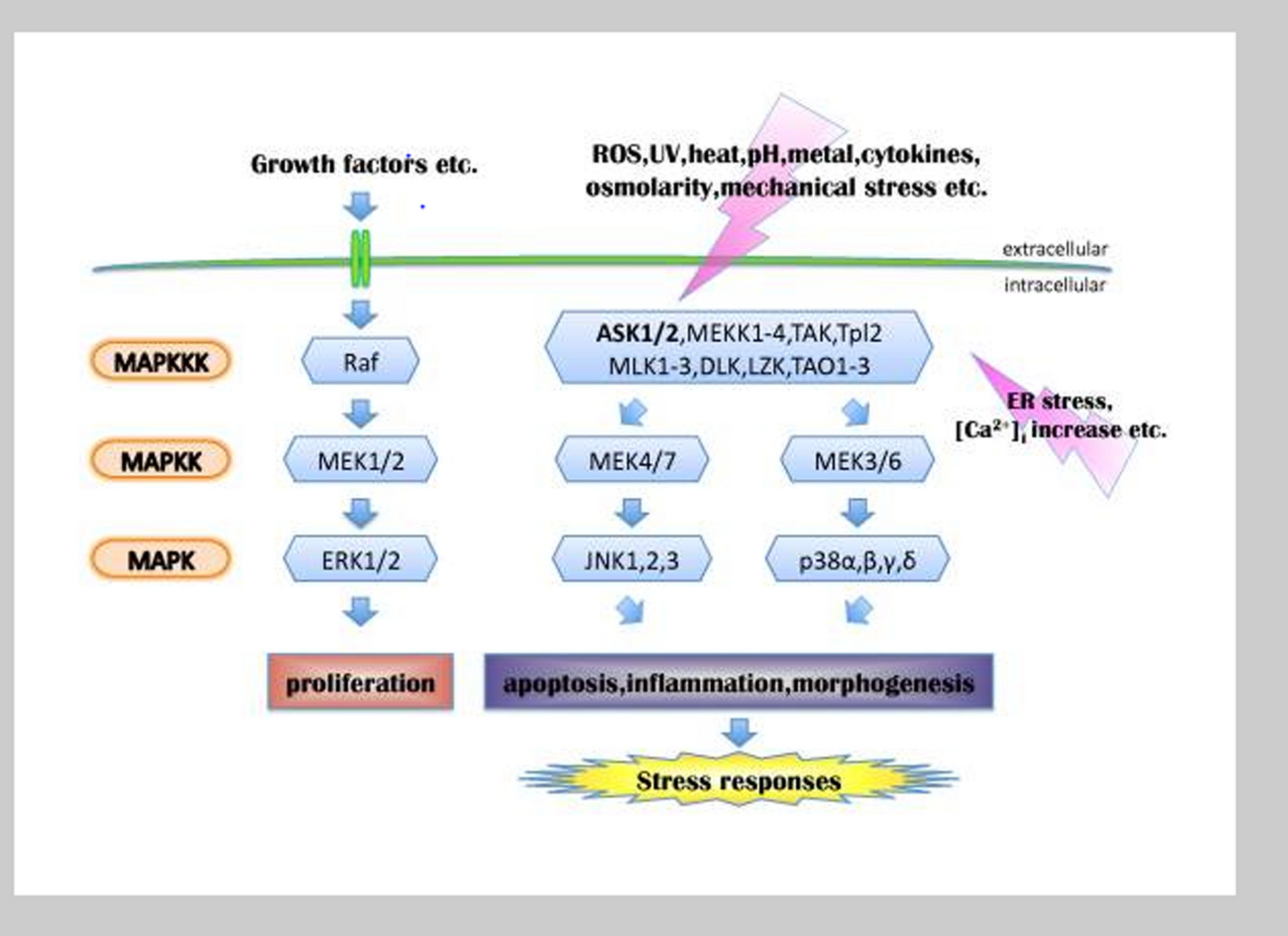
The results showed that SP1 gene expression decreased with the time series and dose in HU-treated cells. In other studies, they have been shown that the effect of some microRNAs, such as miR-23a and miR-27a, on SP1 mRNA, inhibits γ-to-β switching [18, 19]. Moreover, HU inhibits the ERK pathway by activating the MAPK pathway, inhibiting SP1 and fetal hemoglobin increases [20]. The ERK pathways are highly regulated cascades responsible for fundamental cellular processes including cell proliferation and differentiation [21]. The normal maturation of erythrocytes is linked to heightened levels of transcriptional activators GATA-1 and SP1 [22]. GATA-1 plays a significant role in promoting the development of red blood cells by regulating various genes specific to this process, such as globin. Additionally, research has demonstrated that GATA-1 and SP1 collaborate at the γ-globin promoter [22]. Mnika et al. showed that SP1 is a main target of miR-23a, which inhibits switching γ-globin to β-globin. KLF3, a negative regulator of erythrocytes, is also inhibited explicitly by miR-27a [23]. The results of this study are in line with the study of Mnika et al. which showed that HU prevents gamma-to-beta switching by inhibiting SP1.
The results demonstrated that IGF2BP3 gene expression increased after HU treatment. The IGF2BP3 gene is highly expressed in the early stages of development of most tissues and declines after birth. This protein induces erythroblast differentiation and maturation. Also, IGF2BP3 plays an important role in the differentiation process at the early stages of embryogenesis, by binding to three IGF2 leader mRNA and blocking its expression [15]. Jaira et al. showed that induction of IGF2BP1 expression increased γ-globin gene. IGF2BP3 is evolutionarily regulated in human erythroid tissues by being silenced during fetal–adult transition. However, the effect of IGF2BP3 on HbF levels was lower than that of IGF2BP1 in cultured adult erythroblasts [24]. The results of this study are consistent with Jaira et al.’s study that the expression of IGF2BP3, like IGF2BP1, increases γ-globin gene expression [24].
The results demonstrated that COL4A5 gene expression was increased in K562 cells exposed to HU. Hydroxyurea is known to induce the expression of γ-globin by miRNA-mediated silencing of BCL11A, a gene involved in switching from γ to β globin gene expression during erythropoiesis in adulthood. The gene expression modulating effects of HU can be mediated through three main mechanistic actions, including epigenetic modifications, post-transcriptional effects, and signaling mediators, including MAPK, PI3K/Akt, and MYB [25]. In this study, we demonstrated that HU treatment can be associated with the up-regulation of COL4A5 along with the induction of γ-globin expression. It is currently unknown whether a similar HU-mediated regulatory pathway controls the expression of COLA45 and γ-globin, warranting more research in this area. It has been noted that COL4A5 can activate phosphatidylinositol 3-kinase PI3K/protein kinase B (AKT) signaling pathway [26], contributing to the expression of γ-globin [27]. There is a need for further evaluations to divulge other possible roles of COL4A5 in HU-mediated cellular and molecular functions.
The results demonstrate that BCL2 gene expression decreased significantly after 12 h, but increased again at 24 and 48 h. BCL2 proto-oncogene inhibits the apoptosis pathway. HU is a compound that acts on ribonucleoside diphosphate reductase and inhibits DNA synthesis. Caspase-3 activation is an important factor in the activation of the apoptosis cascade. HU increased caspase-3 activity 20 to 30 times higher than the untreated cells. Therefore, there is a correlation with this reagent-induced apoptosis [28]. In a study, it was reported that HU induced apoptotic death of K562 cells through BCL2 protein down-regulation and Bax protein up-regulation and degraded the PARP protein in a p53-independent pathway [29]. The results are consistent with other studies showing that HU induces apoptosis through down-regulation of the anti-apoptotic protein BCL2, inhibiting apoptosis and keeping cells alive.
The findings demonstrated that HU notably increased the expression of the γ-globin gene after 24 and 48 h. Pule et al. found that when HU down-regulated BCL11A, MYB, and KLF-1, it resulted in an increase in γ-globin expression. Their study revealed a connection between the expression patterns of miR-15a, miR16-1, miR-210, miR-29a, miR-96, and the genes MYB, BCL11A, and KLF-1 in response to HU treatment. They proposed a direct interaction between miR-26b and the MYB 3′-UTR as a possible explanation for the up-regulation of γ-globin through the MYB/KLF-1/BCL11A pathway [30]. Consequently, blocking the c-MYB pathway results in an increase of HbF production. According to Barbarani and colleagues, a heightened HbF expression in cultured red blood cells was linked to decreased c-MYB levels [31]. Although these results may help clarify the molecular mechanism of the γ-globin gene switching process, further in vivo and clinical studies are still needed to find alternative therapies for β-thal patients.




Comments (0)