
記住我
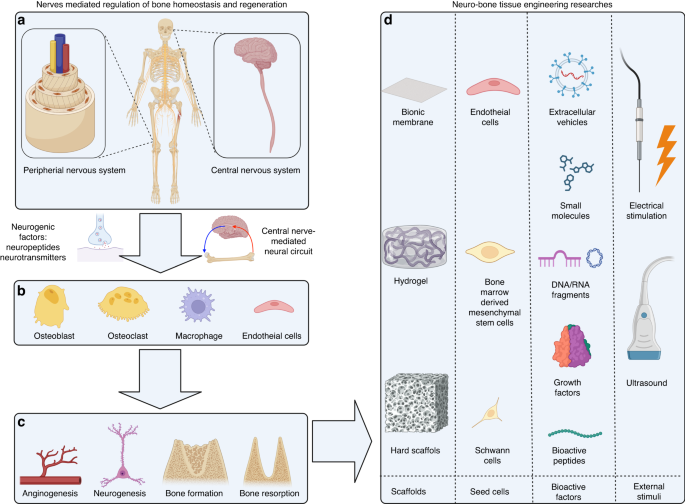


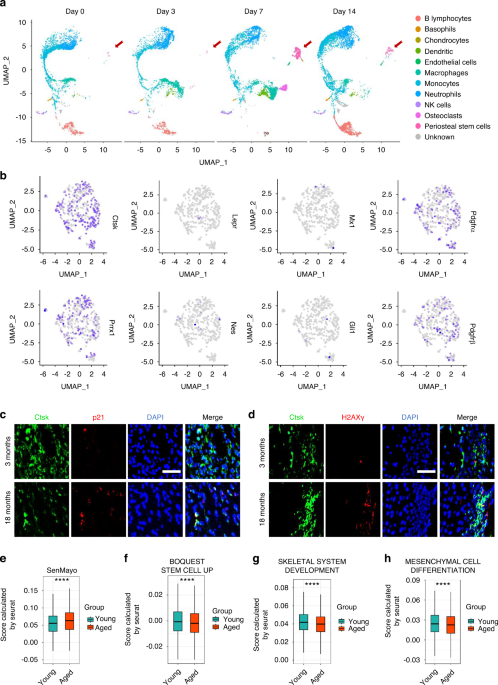
In the osteoblast specific conditional Wwtr1 knockout mouse model, we found a gene-dosage effect on osteoblast-mediated bone formation and bone mass (Figs. 1 and S1). Compared with control mice (Wwtr1+/+), heterozygous conditional Oc-Cre;Wwtr1flox/+ (Wwtr1Oc-Het), which has an approximately 37% reduction in Wwtr1 message expression in bone (Table 1), showed 15% and 12% reductions of BMD in male and female adult mice, respectively (Fig. 1a, b). Oc-Cre;Wwtr1flox/− (Wwtr1Oc-cKO) mice, which had a 64% reduction in Wwtr1 message expression in bone, showed an even greater loss of bone mass, with 24% and 22% reductions of BMD in male and female adult mice, respectively. Micro-CT 3D analysis showed that the reduction in bone mass in single conditional Wwtr1Oc-Het heterozygous mice arose from a 23% reduction in trabecular bone volume and a 11% reduction in cortical bone thickness in both male and female adult mice. Conditional Wwtr1Oc-cKO mice had a 44% loss in trabecular volume and 20% loss in cortical thickness.
Fig. 1
Conditional deletion of Wwtr1 in mature osteoblasts on postnatal bone homeostasis. a, b Bone mineral density by DEXA scan in male and female mice. c Bone structure by micro-CT 3D images analysis from both male and female mice. d Periosteal mineral apposition rate (MAR) by calcein double labeling. There was a significant reduction in periosteal MAR in single Wwtr1Oc-Het heterozygous mice compared with age-matched control mice and an even greater decrement in Wwtr1Oc-cKO null mice, indicating a gene-dosage effect of Wwtr1 on osteoblast-mediated bone formation. e TRAP staining (red color) for osteoclast activity. Data are expressed as the mean ± S.D. from serum samples of individual mice (n = 6). *P < 0.05, **P < 0.01, ***P < 0.001 compared with wild-type control mice. P values were determined by 1-way ANOVA with Tukey’s multiple-comparisons test
Table 1 Gene-expression profiles in bone from Wwtr1-deficient miceIn agreement with the DEXA and micro-CT data, analysis of bone histology showed a Wwtr1 gene-dosage dependent reduction in trabecular bone volume (Figs. 1c and S1a) and cortical bone thickness (Figs. 1c and S1b) in the distal femur and a decrease in bone formation rate measured by double calcein labeling (Fig. 1d). There were 43% and 55% reductions of periosteal MAR in both conditional Wwtr1Oc-Het heterozygous and Wwtr1Oc-cKO null mice, respectively, compared with control mice (Wwtr1+/+). Unexpectedly, conditional deletion of Wwtr1 in osteoblasts resulted in enhanced osteoclast activity, as evidenced by increased TRAP immunostaining in the growth plate of conditional Wwtr1Oc-cKO null mice (Fig. 1e).
Real-time RT-PCR analysis revealed a Wwtr1 gene-dosage dependent changes in osteoblast markers including Runx2-II and Wnt10b, FGF-23, Mepe, RankL, CYR61, and CTGF and chondrocyte markers such as Collagen II and Collagen X. Reductions of Runx2-II and Wnt10b impairs osteoblast proliferation and differentiation. Increments of FGF-23 and Mepe as well as Yap signaling such as increased CYR61 and CTGF gene expressions inhibits osteoblast differentiation and mineralization. An increase in RankL and the RankL/OPG ratio promotes osteoclast differentiation, leading to greater TRAP staining and higher osteoclast activity in conditional Wwtr1Oc-cKO null mice compared with control mice (Wwtr1+/+). Conditional deletion of Wwtr1 also resulted in increased adipocyte markers such as PPARγ2, aP2 and Lpl gene expressions (Table 1).
Unexpectedly, global Wwtr1+/− heterozygous mice, which had a ~40% reduction in Wwtr1 message expression in bone, did not have significant changes in BMD or bone volume. Single-heterozygous Wwtr1+/– showed normal bone gene expression profiles as well (Table 1). The osteoblast specific reduction but not global reductions of Wwtr1 on the skeletal phenotype points to other important co-factors in osteoblasts, such as Wwtr1 interactions with PC1, or counteracting effects of Wwtr1 in non-osteoblastic cells.
Additive effects of combined Pkd1 and Wwtr1 deficiency to reduce bone massTo test the functional effects of the interaction between Pkd1 and Wwtr1 in osteoblasts, we characterized Osteocalcin-Cre;Pkd1flox/−; Wwtr1flox/− (Pkd1/Wwtr1Oc-cKO) double null mice with osteoblast-specific conditional deletions of both Pkd1 and Wwtr1 in bone. In the conditional Pkd1/ Wwtr1 double knockout model, we found that Pkd1 and Wwtr1 have additive effects in reducing osteoblast-mediated bone formation. Compared with control mice (Pkd1flox/+;Wwtr1flox/+), both conditional Pkd1Oc-cKO and Wwtr1Oc-cKO null mice had similar reductions in bone mass, as evidenced by respective 22% and 21% reductions of BMD (Fig. 2a, b), 33% and 35% reductions in trabecular bone volume (Figs. 2c and S2a), and 18% and 19% reductions in cortical bone thickness (Figs. 2c and S2b) as determined by distal femur Micro-CT 3D image analysis and Goldner staining. Also, the reductions in bone mass were similar in male and female adult mice (Fig. 2a, b).
Fig. 2
Conditional deletion of Pkd1 and Wwtr1 in mature osteoblasts on postnatal bone homeostasis. a, b Bone mineral density by DEXA scan in male and female mice. c Bone structure by micro-CT 3D images analysis from male mice. d Periosteal mineral apposition rate (MAR) by Calcein double labeling. There was a significant reduction in periosteal MAR in single Pkd1Oc-cKO or TAZOc-cKO mice compared with age-matched control mice and an even greater decrement in double Pkd1/TAZOc-cKO null mice, indicating an additive effect of PC1 and TAZ on osteoblast-mediated bone formation. e TRAP staining (red color) for osteoclast activity. Data are expressed as the mean ± S.D. from serum samples of individual mice (n = 6). *P < 0.05, **P < 0.01, ***P < 0.001 compared with wild-type mice, #P < 0.05, ##P < 0.01, ###P < 0.001 compared with Wwtr1Oc-cKO mice, and &P < 0.05, &&P < 0.01, &&&P < 0.001 compared with Pkd1Oc-cKO mice, respectively. P values were determined by 1-way ANOVA with Tukey’s multiple-comparisons test
The skeletal phenotype of double Pkd1/Wwtr1Oc-cKO mice was more severe than either single Pkd1Oc-cKO or Wwtr1Oc-cKO null mice. Double Pkd1/Wwtr1Oc-cKO mice had greater losses in BMD, trabecular bone volume, and cortical bone thickness with 33%, 53%, and 27% reductions, respectively in both male and female adult mice. This indicates the additive effects of Pkd1 and Wwtr1 in postnatal bone homeostasis (Figs. 2 and S2). Consistent with lower bone mass, combined Pkd1 and Wwtr1 deficiency also resulted in additive reductions in osteoblast-related gene expressions, such as in Runx2-II, Osteocalcin, and Dmp1 (Table 2), as well as mechanosensing responsive genes such as in Wnt10b, c-Jun, and PTGS2 (Table 2). Periosteal MAR (Fig. 2d) was decreased by ~73% in conditional double Pkd1/Wwtr1Oc-cKO mice compared to controls, whereas Wwtr1Oc-cKO and Pkd1Oc-cKO single conditional knockout mice had reductions in periosteal MAR of 55% and 53%, respectively compared to control mice. Loss of either Pkd1 or Wwtr1 resulted in enhanced marrow adipogenesis, but no additive effects on adipocyte differentiation-related gene expressions were observed in the conditional double Pkd1/Wwtr1Oc-cKO mice (Table 2).
Table 2 Gene-expression profiles in bone from Pkd1Oc-cKO or/and Wwtr1Oc-cKO miceWe found that the conditional deletion of Pkd1 or Wwtr1 in osteoblasts has opposite effects on osteoclast activities. There was a decreased RankL/OPG expression ratio and TRAP immunostaining in Pkd1Oc-cKO mice but an increased RankL/OPG expression ratio and TRAP immunostaining in Wwtr1Oc-cKO mice (Table 2 and Fig. 2e). In contrast, double Pkd1/Wwtr1Oc-cKO had similar RankL expression and TRAP immunostaining when compared to control, indicating a recovery of osteoclast activities in the double null mice (Table 2 and Fig. 2e). Changes in gene expression and immunostaining in bone correlated with alterations in serum biomarkers (Table 3). In this regard, further evidence for osteoblast and osteocyte dysfunction includes reductions in serum osteocalcin and FGF-23 from in single Pkd1Oc-cKO or Wwtr1Oc-cKO mice compared with age-matched control mice and an even greater decrement in double Pkd1/Wwtr1Oc-cKO null mice (Table 3). In contrast, serum levels of TRAP, a marker of bone resorption, were decreased in single Pkd1Oc-cKO mice, increased in single Wwtr1Oc-cKO mice, but restored in double Pkd1/Wwtr1Oc-cKO null mice compared with control littermates (Table 3). In addition, serum level of leptin was significantly higher in single Pkd1Oc-cKO or Wwtr1Oc-cKO mice than age-matched control mice. However, we did not observe further increase in double Pkd1/Wwtr1Oc-cKO null mice (Table 3). These findings suggest that Pkd1 and Wwtr1 have distinct functions among osteoblasts, adipocytes, and osteoclasts in bone in vivo.
Table 3 Biochemistry analysis of serum from Pkd1Oc-cKO or/and Wwtr1Oc-cKO miceLoss of mechanical loading response in conditional Pkd1 and Wwtr1 deficient miceThe cross-sections of tibiae from control and double Pkd1/Wwtr1Oc-cKO null mice after mechanical tibia loading studies in vivo are shown in Fig. 3. In wild-type control mice, loaded tibia showed a 2-fold increase in periosteal mineral apposition rate. In contrast, there was no measurable increase in periosteal mineral apposition in the loaded tibia from double Pkd1/Wwtr1 knockout mice (Fig. 3). In addition, a real-time RT-PCR analysis revealed that loaded tibia from the control mice had a dramatic response to mechanical stimulation, evidenced by significant increases of mechanosensing (e.g., Wnt10b, FzD2, Axin2, PTGS2, c-Jun, c-Fos, and Runx2-II) and osteogenic (e.g., Osteocalcin, Alp, Collagen I, and Dmp1) but decreases of adipogenic (e.g., PPARγ2, aP2, and Lpl) gene expressions when compared with no load control tibia. In contrast, even when using the same loading regimen, no changes of mechanosensing, osteogenic, and adipogenic gene expression profiles were observed in the loaded tibia from double Pkd1/ Wwtr1Oc-cKO null mice when compared with no load control tibia (Table 4). Thus, PC1 and Wwtr1 are important in mediating mechanotransduction in bone.
Fig. 3
An impairment of anabolic response to mechanical loading in conditional Pkd1 and Wwtr1 deletion in bone. Representative images of midshaft tibia cross sections from no-load and loaded ulnae of wild-type control and compound Pkd1/Wwtr1Oc-cKO null mice after loading. Data are mean ± S.D. from 6 tibias of wild-type control and Pkd1/Wwtr1Oc-cKO mice. ***P < 0.001 compared with wild-type mice. P values were determined by 1-way ANOVA with Tukey’s multiple-comparisons test
Table 4 Mechanosensing, osteogenic, and adipogenic gene-expression profiles that respond to mechanical loading in wild-type and double Pkd1/Wwtr1Oc-cKO miceValidation of MS2 key binding to residues in PC1/PC2 C-terminus tailsWe have previously showed that the small molecule MS2 activates PC1/PC2 complex signaling.2 Using computational modeling, we engaged in an induced fit docking campaign and predicted several potential ligand binding complexes. From these predicted poses, we identified key residues in the PC1 and PC2 C-terminus tail regions with which MS2 is predicted to interact. As shown in Fig. 4, the PC2-CTT binding site for MS2 is predicted to include Lysine874 and Arginine877, whereas the PC1-CTT binding site for MS2 involves Tyrosine4236 (Fig. 4a, b). To test these predictions, we performed site-mutagenesis of key residues in both PC1 and PC2 and tested the effects of MS2 on PC1 and PC2 assembly using a BRET2 assay. We observed that the compound MS2 markedly enhances the BRET2 signal in wild-type constructs, while mutagenesis of key residues in either PC1-CTT or PC2-CTT constructs completely abolished the BRET2 signal in the presence of compound MS2, confirming the role of MS2 in enhancing PC1 and PC2 interactions and lending support that MS2 is binding to the protein complex (Fig. 4c, d).
Fig. 4
Cell-based BRET2 assays for MS2-target engagement assays. a Chemical structure of MS2 and an example of predicted 3D binding mode for MS2 (ball-and-sticks rendering in CPK colors) in PC2 (light blue). PC1 (red) as bound to PC2 in the homology structure is superimposed. b An example of calculated 2D binding mode and residues for MS2 in PC1/PC2 C-tails. c A diagram of BRET2 constructs and reactions in the presence of DeepBlue C with or without MS2 stimulation. d BRET2 signal changes from wild-type and mutant constructs with or without MS2 incubation. e, f Time-dependent changes of PC1 and PC2 proteins with or without MS2 incubation during osteogenic differentiation cultures in MC3T3-E1 cell line. Incubation Data are presented as the mean ± SD from 3 independent experiments (n = 3). **P < 0.01 compared with vehicle control
Next, we examined PC1/PC2 complex formation during MC3T3-E1 osteoblast differentiation in vitro. We observed culture duration dependent increase of PC1/PC2 complex formation by western blot analysis. Incubation with 1 μmol·L−1 of MS2 in osteogenic cultures markedly increased the amount of PC1 and PC2 protein as assessed by western blot analysis (Fig. 4e, f). These data suggests that MS2 may molecularly stabilize the PC1/PC2 complex in osteoblast culture in vitro.
Loss of MS2-mediated stimulated increase in bone mass in conditional Pkd1 and Wwtr1 deficiency miceFinally, we treated wild-type and conditional double Pkd1/Wwtr1Oc-cKO null mice with vehicle or MS2 (50 mg·kg−1) i.p. daily and assessed their skeletal response. After only 2 weeks, we observed that wild-type control mice treated with MS2 had a 15% increment in femoral bone mineral density compared to vehicle control (Fig. 5). Micro-CT 3D images revealed that MS2 treated wild-type mice had a 39% increase in trabecular bone volume and 16% increase in cortical bone thickness.
Fig. 5
Effects of MS2 on bone formation in wild-type and compound Pkd1/Wwtr1Oc-cKO null mice. a Bone mineral density by DEXA scan. b Bone structure by micro-CT 3D images analysis. c Periosteal mineral apposition rate (MAR) by Calcein double labeling. d Osteoclast activities by TRAP immunostaining after MS2 (50 mg·kg−1) treatment for 4 weeks compared to vehicle control. Data are mean ± S.D. from 6 tibias of wild-type control and compound Pkd1/Wwtr1Oc-cKO null mice. *P < 0.05, **P < 0.01, ***P < 0.001 compared with wild-type control mice. P values were determined by 1-way ANOVA with Tukey’s multiple-comparisons test
In contrast, administration of MS2 had no effects on bone mineral density and bone structure in double Pkd1/Wwtr1Oc-cKO null mice, suggesting specific-target dependent effects of MS2 on polycystins/Wwtr1 signaling (Fig. 5). We also observed that there were 1.6-fold increases in bone formation rate in wild-type mice treated with MS2 compared to the vehicle control, in agreement with enhanced osteoblastogenesis (e.g., Runx2-II, Osteocalcin, ALP and Dmp1) and suppressed marrow adipogenesis (e.g., PPARγ2, aP2, and Lpl) by a real-time RT-PCR analysis (Table 5 and Fig. 5). Again, administration of MS2 had no effects on bone formation rate and bone gene expression profiles in double Pkd1/Wwtr1Oc-cKO null mice. Furthermore, MS2 stimulated mechanosensing gene expressions, including Wnt1, Wnt10b, Axin-2, FzD2, c-Jun, c-Fos, eNOS, and PTGS2, consistent with MS2 acting as a small molecule “mechanomimetic”. MS2 treatment decreased RankL/OPG expression ratio and TRAP immunostained osteoclasts in the MS2-treated mice compared to vehicle control (Table 5 and Fig. 5). Administration of MS2 had no effects on osteoblast-mediated bone formation rate, marrow adipogenesis, and osteoclast activity in conditional double Pkd1/Wwtr1Oc-cKO null mice (Table 5 and Fig. 5). These data support that MS2 functions as anabolic drugs through the polycystins/Wwtr1 pathway to promote the bone remodeling process.
Table 5 Gene expression profiles in bone from MS2-treated wild-type control and Pkd1/Wwtr1Oc-cKO mice
留言 (0)