
記住我
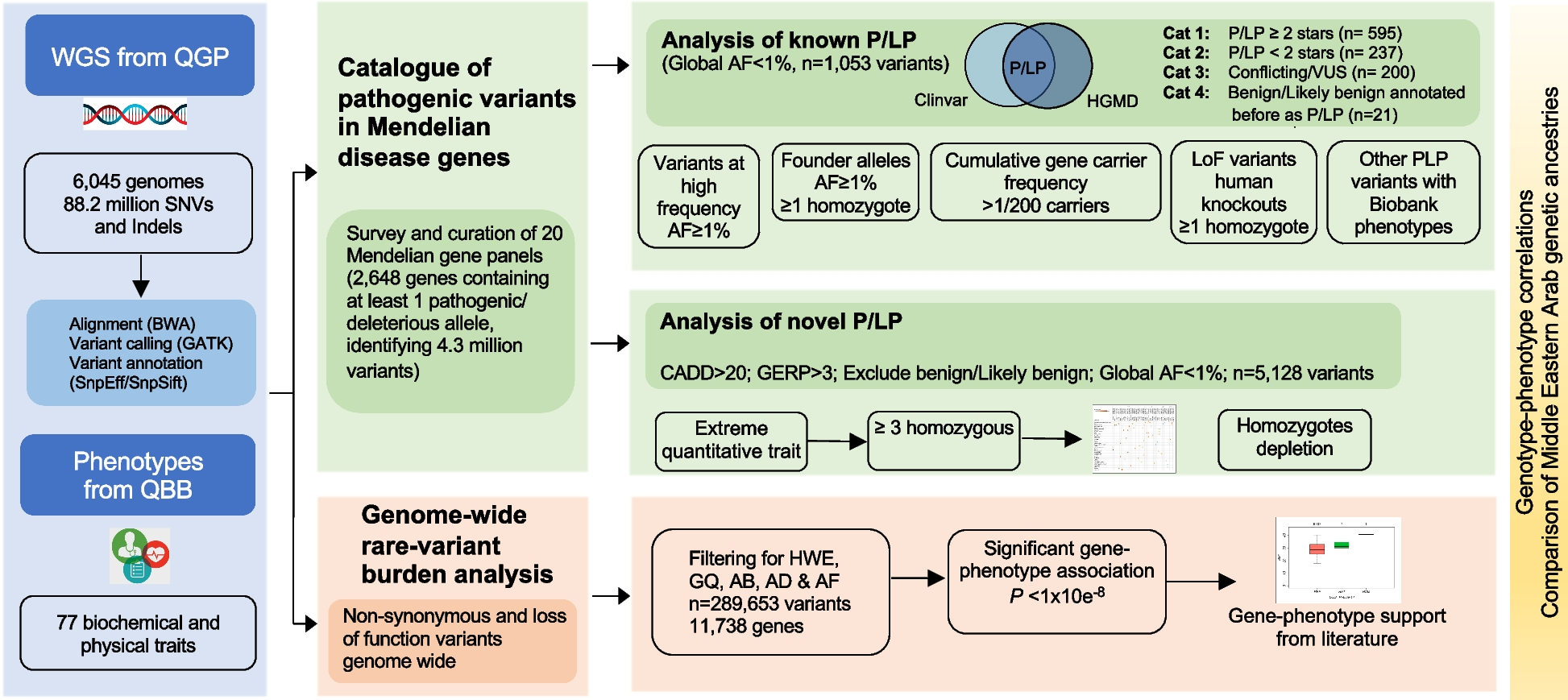





To induce mouse liver tumors using multiplexed CRISPR/Cas9, we chose 34 TSGs along with a 5′-upstream site of the Setd5 locus as a negative control for CRISPR/Cas9-mediated gene inactivation (Fig. 1A; Additional file 2: Fig. S1). These 34 TSGs are involved in at least 11 cancer-related signaling pathways (Fig. 1A). We designed and constructed at least 3 sgRNAs for each target gene except p53 and analyzed these sgRNAs for their efficiency of frame-shift editing using our BGN reporter established previously (Additional file 2: Fig. S2A) [43]. The 23-bp target sequence containing the PAM for each sgRNA was inserted into the I-SceI-EcoRI site of the BGN reporter (Additional file 2: Fig. S2A). To test each sgRNA, the expression plasmids for Cas9 and sgRNA and the BGN reporter plasmid containing the Cas9-sgRNA target were transfected into mouse embryonic fibroblast cell line NIH-3T3 cells and the frequencies of “3n + 1”-bp frame-shift measured by flow cytometry for GFP+ cells (Additional file 2: Fig. S2B). After testing of each sgRNA, we selected the most effective sgRNA for each target gene to establish a plasmid library of 35 sgRNAs. We then used hydrodynamic tail vein injection (HTVI) to deliver this sgRNA plasmid library mixed with the Streptococcus pyogenes Cas9 (SpCas9) expression plasmid into mouse liver cells to induce liver tumors [19, 48]. Total volume injected was 2 mL with the amount of the SpCas9 plasmid fixed at 200 μg for each mouse. The amounts of each sgRNA in the sgRNA library injected ranged from 0.005 μg to 5 μg (Fig. 1B). Liver tumors were formed and visible in mice within 30–60 days (Fig. 1B–C). Dilution of sgRNA delayed development of Cas9/sgRNA-induced mouse liver cancer and reduced cancer induction (Fig. 1B). After each sgRNA was reduced to 0.2 μg, tumor development rarely occurred (Fig. 1B). Histological analysis of tumor nodules indicated that the hepatic lobule structures were destructed in tumor nodules (Fig. 1D). Immunostaining for the HCC biomarkers AFP and GP73 and the ICC biomarker CK19 revealed the presence of HCC, ICC, and mixed HCC-ICC types in mouse liver tumors induced by CRISPR/Cas9 (Fig. 1D). In one section of a single tumor nodule, three selected regions exhibited distinct histologic features (Fig. 1E), indicating strong ITH in CRISPR/Cas9-induced liver cancer in mice.
Fig. 1
Induction of primary mouse liver cancer by CRISPR/Cas9-mediated somatic mutagenesis in mice. A 34 TSGs and the Setd5 control site targeted by multiplexed CRISPR/Cas9. Each TSG is indicated with its related cancer signaling pathway. Hepatic delivery of SpCas9 and sgRNA expression plasmids is achieved by hydrodynamic tail vein injection (HTVI). B Effect of sgRNA dosage on CRISPR/Cas9-induced liver tumor formation. Each dot indicates one mouse. C Mouse liver specimen with or without tumor nodules (left) and H&E staining of a tumor nodule (right). D Microscopic IHC images of CRISPR/Cas9-induced liver tumors. T#1, T#2, and T#3 represent tumor nodules from three mice. AFP and GP73: HCC biomarkers; CK19: ICC biomarker. E Histology and IHC staining of a liver tumor nodule indicating three heterogeneous regions. Left: these three regions indicated with R1, R2, and R3 in H&E staining; Right: microscopic IHC images for these three regions
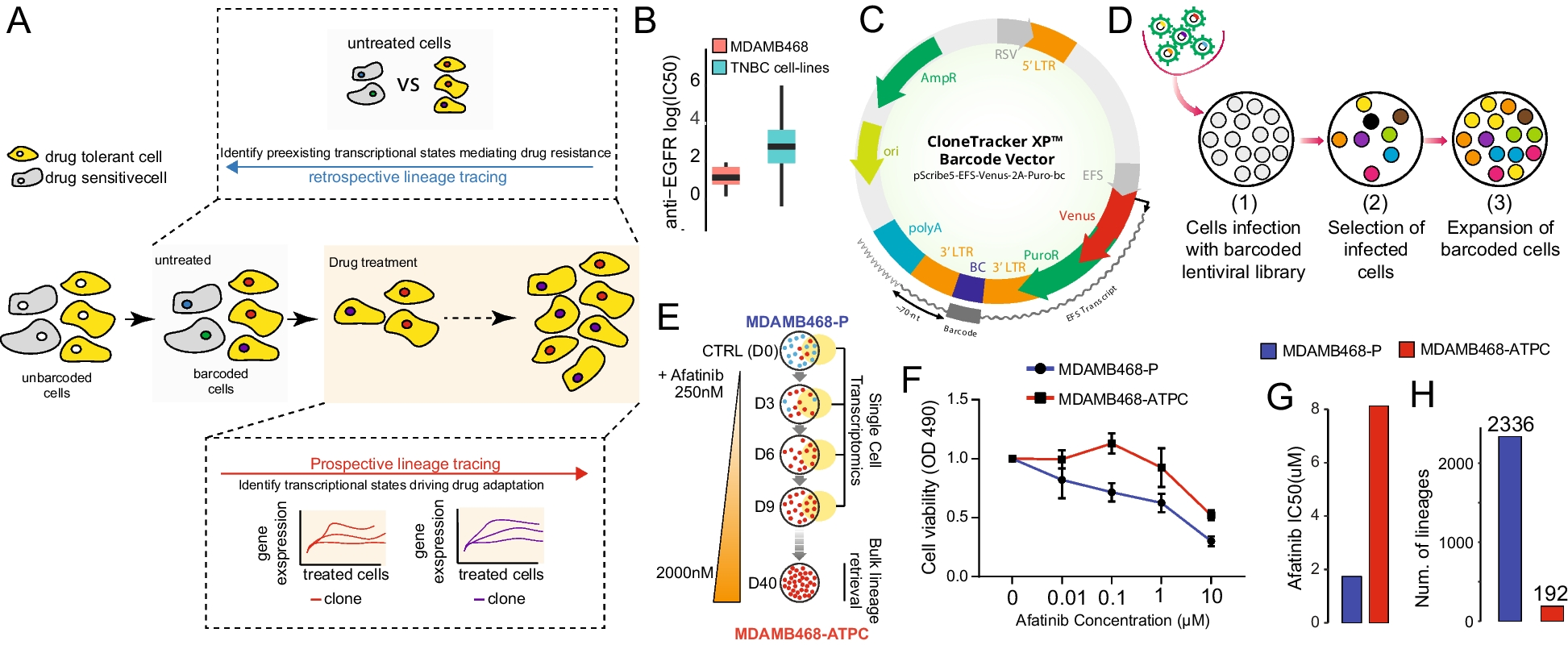
Heterogeneity of targeted TSG mutations in CRISPR/Cas9-induced mouse liver tumorsWe performed next generation sequencing (NGS) of PCR-amplified target sites in tumor nodules to identify targeted TSG mutations and determine the frequencies and spectra of these mutations. We define an allelic mutation with a frequency at no less than 5% as a true mutation to reduce the interference of sequencing errors. While different targets showed different mutational spectra, we also found that many same target sites carried different allelic mutations with varying frequencies. For example, in analysis of a tumor nodule induced by SpCas9 together with 35 sgRNAs each at 0.8 mg, in addition to their respective wild-type (WT) allele, the p53 target site had 3 types of mutations, 28.12% for 1-bp deletion on the left side of the break (termed Del1|0), 16.13% for 7-bp deletion on the right side of the break (Del0|7), and 13.87% for insertion of 1A at the break (Ins1A); Atm had two (23.81% for D6|0 and 20.07% for Ins1C), and Rb1 had none in addition to WT alleles (Fig. 2A). In this tumor module, majority of target genes were mutated at intended sites and the mutation frequencies (MF) were high at some of these sites after the frequencies of all allelic mutations were combined for each site (Fig. 2B). This was further confirmed by analysis of more tumors induced by SpCas9 together with 35 sgRNAs each at 1 or 3 mg (Fig. 2C; Additional file 1: Table S2; Additional file 2: Fig. S3). The mutation profiles differed between tumors within a same mouse or from different mice, indicating intertumoral heterogeneity in CRISPR/Cas9-induced liver cancer with respect to targeted mutations of TSGs.
Fig. 2
Target TSG mutation profiles in CRISPR/Cas9-induced mouse liver tumor nodules and single-cell clones. A Representative target mutation types of p53, Atm and Rb1 in a tumor nodule induced by SpCas9 together with 35 sgRNAs each at 0.8 mg and the frequencies of these mutations. sgRNA target sites with PAM and the cleavage position are indicated. Dash lines and nucleotides in curly bracket in the sequences denote deletion and inserted nucleotides, respectively. WT, wild-type; Del, deletion; Ins, insertion. The numbers flanking Del or Ins indicate the numbers of deleted or inserted nucleotides. B Representative target mutations of 21 TSGs with cumulative frequencies in the tumor nodule in A. Fourteen unedited gene are also listed on the right. C Mutational landscape of 35 targets in 22 primary tumor nodules, 4 parts from a single tumor nodule, and 37 single-cell clones as indicated. Top and right bars indicate the mean of the mutation frequencies for each sample and each gene, respectively. Cumulative matrix mutation frequencies are highlighted from the highest 8 in red to the lowest 0 in blue
In addition, a tumor nodule from mouse #5 was microdissected into four regions, and each region analyzed for targeted TSG mutations by NGS of PCR-amplified target sites. Significant variations existed in mutation profiles between these four regions (Fig. 2C; Additional file 1: Table S2; Additional file 2: Fig. S3), suggesting ITH of targeted TSG mutations. These variations may be also attributable to varying healthy stromal components and/or residual normal tissue in individual tumor nodules.
To further analyze ITH of targeted TSG mutations, we established single-cell clones from tumor nodules 5T2, 6T1, 7T1, and 8T1 from different mice and analyzed targeted mutations in these tumor cells by targeted PCR amplicon deep sequencing (Additional file 2: Fig. S3). Intriguingly, approximately half of 34 TSGs were highly mutated between individual clones from the same nodules (in red in Fig. 2C). For some other targeted genes, e.g., Smad4 and Tsc1, the frequencies of targeted mutations varied widely between 0 and 100% among single-cell clones from the same nodules, e.g., 1T1 (Fig. 2C). Between different clones from the same nodules, some targets sites carried identical mutations with similar frequencies but some harbored identical mutations with different frequencies or even different mutations (Fig. 2C; Additional file 1: Table S3). These results indicated strong ITH of targeted TSG mutations in CRISPR/Cas9-induced mouse liver cancer and suggested that single-cell clones originating from the same nodules undergo multiple edits by this 34-sgRNA library of TSGs.
As observed previously [22], individual target sites often carried more than two different allelic mutations within a tumor nodule (Additional file 1: Table S2). This could be explained at least by the following three possibilities. (1) Several transfected founder cells happened to start together with a different subset of mutations induced by multiplexed CRISPR/Cas9 genome editing and developed into a single tumor nodule. (2) Some mutations were first induced in a single transfected founder cell by multiplexed CRISPR/Cas9 genome editing and others occurred only after the first cell division in subsequent daughter or granddaughter cells [22]. (3) More than two copies of target sites exist in the genome of a single transfected founder liver cell due to polyploidization of liver cells [49,50,51] and are differently mutated by CRISPR/Cas9. However, in some single-cell clones (e.g., 1C3 and 6C7), a few target sites harbored more than two different allelic mutations and even up to 9 different allelic mutations, some of which occurred with varying frequencies (Additional file 1: Table S3). This was unexpected for a single-cell clone where only two copies of a target site are the most likely for CRISPR/Cas9-induced mutations unless the cells are hyperploid or the target sites have repeat sequences. Nevertheless, this type of copy number variations (CNVs) at the single-cell level represents a new source for heterogeneity of targeted TSG mutations in CRISPR/Cas9-induced mouse liver cancer.
Type and frequency of targeted mutation alterations from parental clones to subclonesGiven the presence of more than two allelic mutations with significant different frequencies in a single-cell clone, we wondered whether this mutation pattern is inheritable. We thus selected two single-cell clones (i.e., 1C3 and 6C7) that carried more than two copies of allelic mutations at some target sites, isolated 4 subclones from 1C3 and 8 subclones from 6C7, and analyzed targeted mutations in these clones and subclones. The profile of target site mutations was different between parental clones and their respective subclones and between subclones (Fig. 3A; Additional file 1: Table S4). For instance, 6 Rb1 target site variants, 8 Lkb1 target site variants, 4 Arid1a target site variants, and 5 Smad4 target site variants were detected across parental 1C3 clone and its 4 subclones, but each type displayed different frequencies within individual clones and the mutation profiles were also different between clones (Fig. 3B–E). The frequency of the wide type Rb1 was less than 2% in paternal clone 1C3 but over 30% in 4 subclones (Fig. 3B). In contrast, the frequency of the Rb1 Del22|5 mutation was nearly 30% in 1C3 but hardly detectable in subclone 1C3-1 or 1C3-2, less than 3% in 1C3-3 and about 5% in 1C3-4 (Fig. 3B). The Lkb1 Del4|0 mutation was barely detected in 1C3, but the frequencies of this mutation were 20% or more in 4 subclones (Fig. 3C). The Lkb1 Del10|0 mutation was a dominant mutation with the frequency at over 30% in 1C3 and its subclone 1C3-4 but less than 1% in subclone 1C3-1 and 1C3-2 and around 16% in subclone 1C3-3 (Fig. 3C). The Arid1a Del11|16 mutation was dominant at more than 70% in 1C3-1 but negligible at less than 2% in 1C3-4 (Fig. 3D). In contrast to Del11|16, the Arid1a Del10|9 mutation was nearly undetectable in 1C3-1 but highly frequent at more than 50% in 1C3-4 (Fig. 3D). Similarly, while the Smad4 WT allele was detected at over 60% in 1C3 but less than 6% in 1C3-4, the frequency of the Smad4 Del2|0 was 2% in 1C3 but nearly 35% in 1C3-4 (Fig. 3E).
Fig. 3
Alteration of target site mutations between subclones of single-cell clone 1C3 and 6C7 derived from mouse liver tumors. A Changes in target site mutation profiles between parental single-cell clones (i.e., 1C3 and 6C7) and their subclones (1C3-1, 1C3-2, 1C3-3, and 1C3-4 for 1C3 and 6C7-1, 6C7-2, 6C7-3, 6C7-4, 6C7-5, 6C7-6, 6C7-7, and 6C7-8 for 6C7). Gene targets and their mutation types are shown on top and at bottom, respectively. The numbers flanking Del or Ins indicated at bottom are the numbers of deleted or inserted nucleotides. √Frequency representing square root of mutation frequencies is indicated from the highest 1 in dark green to the lowest 0 in white. The top line chart and the boxplot under show the number of single-cell clones with a specific mutation type, i.e., number of clones, and the percentage distribution of a specific mutation type in all single-cell clones, i.e., proportion (%), respectively. B–E Representative target mutation oscillation of Rb1 (B), Lkb1 (C), Arid1a (D), and Smad4 (E) between parental clone 1C3 and its subclones
The 6C7 parental clone and its subclones were more similar to each other than 1C3 the 1C3 parental clone and its subclones to each other (Fig. 3A; Additional file 1: Table S4). Still, while the Rb1 WT allele in 6C7-2 was over 15%, the other 6C7 subclones as well as the parental clone carried infrequent WT allele of Rb1 (Additional file 2: Fig. S4A). Similarly, the frequency of Lkb1 Del1|0 mutation in 6C7 consistently matched that of its subclones, except for 6C7-1, which displayed a higher mutation frequency (Additional file 2: Fig. S4B). Additionally, the frequencies of three Arid1a target mutation variants oscillated among the 6C7 parental clone and its subclones (Additional file 2: Fig. S4C) and a change in the Smad4 Del2|0 and Del1|0 frequency were observed in 6C7-7 as compared to the 6C7 parental clone and the other subclones (Additional file 2: Fig. S4D). These results together indicate continuing oscillation in the frequencies of target site mutations between these parental clones and subclones. In particular, predominant mutations in parental clones could disappear in subclones whereas negligible mutations in parental clones could appear in subclones.
As spontaneous mutations at a given site normally occur with extremely low probability during cell proliferation [52], it is surprising that the frequency of a specific targeted mutation oscillates significantly from parental single-cell clones to daughter single-cell subclones. We speculated that Cas9-sgRNA might be stably expressed in clones isolated and continue to edit the WT target sites, thus altering the frequency of the targeted mutation. However, we did not detect SpCas9 proteins in the parental clones 1C3 and 6C7 and their subclones (Additional file 2: Fig. S5A). It remains possible that a small amount of SpCas9 proteins stably synthesized in the cells could actively mutate their target sites, even if the protein level is hardly detectable by Western blot. We thus transfected these parental clones or subclones with the expression plasmids for two Col1a1 sgRNAs and one Rosa26 sgRNA with or without the SpCas9 expression plasmid and measured the editing frequencies at the target sites by PCR amplicon deep sequencing. While the editing is efficient with transfection of both SpCas9 and sgRNA, no editing was detected with sgRNA transfection alone or with neither SpCas9 nor sgRNA (Additional file 2: Fig. S5B-C). This excludes the possibility that continuing CRISPR/Cas9 genome editing alters the types and frequencies of targeted TSG mutations from parental clones to subclones.
Continuing type and frequency oscillation of targeted mutations during clonal expansionNext, we asked whether the types and frequencies of targeted mutations change over time during expansion of a subclone. We continuously cultured 4 subclones (i.e., 1C3-1, 1C3-2, 1C3-3 and 1C3-4) and analyzed target site mutations of each subclone on day 0, day 15, and day 30 (Fig. 4A; Additional file 2: Fig. S6). The frequencies of some targeted mutation changed over time during clonal expansion, e.g., from 2.47% at day 0 to 35.09% at day 15 and 34.09% at day 30 for Rb1 Ins1T in 1C3-1 (Additional file 1: Table S5; Additional file 2: Fig. S6). After storage in liquid nitrogen for a year, we thawed and continued to culture the 1C3-1 clone. We repeated analysis of target site mutations in the cultured cells on day 0 (i.e., Mon12), day 90 (i.e., Mon15), and day 180 (i.e., Mon18) (Fig. 4A). The frequencies of some targeted mutations continued to change over time during cell culturing of 1C3-1 (Fig. 4B; Additional file 1: Table S5). For example, the Rb1 Ins1T mutation was negligible on day 0 but appeared with the frequencies at over 30% on day 15 and day 30. Del1|0 mutation was significant at over 60% on day 0 and decreased to about 30% on day 15, day 30, and Mon12 and even 10% on Mon15. On Mon18, this mutation was slightly increased to over 20% (Fig. 4C). Both Lkb1 Del5|2 and Ins1G mutations remained frequent at over 40% from day 0 to Mon12 but decreased to about 20% on Mon15 and increased again to 30% on Mon18 (Fig. 4D). Arid1a Del11|16 and Del10|9 mutations changed in opposite direction on Mon15 and Mon18 (Fig. 4E). Similarly, Smad4 WT and Del16|0 mutations oscillated in opposite direction on Mon15 and Mon18 (Fig. 4F). This data further indicates that some targeted mutations are not stable during proliferation of single tumor cells derived from CRISPR/Cas9-induced liver cancer in mice.
Fig. 4
Oscillation of target site mutations during proliferation of the single-cell subclone 1C3-1 derived from 1C3. A Schematic of experimental outline. Proliferation of the single-cell clone 1C3-1 was divided the first culture cycle that continues for 30 days and the second culture cycle that continues after 1 year storage. Target site mutations were determined by targeted PCR amplicon deep sequencing of gDNA extracted from cells at different time points of cell proliferation as indicated. D0, day 0; D15, day 15; D30, day 30; Mon12, year 1; Mon15, year 1 + 3 months; Mon12, year 1 + 6 months. B Changes in target site mutation profiles of 1C3-1 at indicated time points of cell proliferation (i.e., days 0, 15, and 30 and Mon12, Mon15, and Mon18). Gene targets and their mutation types are shown on top and at bottom, respectively. The numbers flanking Del or Ins indicated at bottom are the numbers of deleted or inserted nucleotides. √Frequency representing square root of mutation frequencies is indicated from the highest 1 in dark green to the lowest 0 in white. The top line chart and the boxplot under show the number of single-cell clones with a specific mutation type, i.e., number of clones, and the percentage distribution of a specific mutation type in all single-cell clones, i.e., proportion (%), respectively. C–F Representative target mutation oscillation of Rb1 (C), Lkb1 (D), Arid1a (E), and Smad4 (F) during cell proliferation of 1C3-1
Type and frequency alteration of targeted mutations from single-cell clones to subcutaneous graftsIn order to evaluate this instability of targeted TSG mutations in vivo, we implanted the subclone 1C3-1 cells into 4 immunodeficient SCID mice subcutaneously (Fig. 5A). All mice grew a visible tumor in about 5 days. We harvested the tumor tissues from these 4 mice at the 14th day and established 2 single tumor cell clones from each tumor tissue (Fig. 5A). Analysis of targeted mutations revealed that tumor tissues harbored more frequent WT allele than both 1C3-1 and tumor cell clones derived from tumor tissues, likely due to the presence of normal cells in subcutaneous tumor samples (Additional file 1: Table S6). Therefore, in order to exclude the interference of normal cells from tumor tissues on the frequency of targeted mutations in tumor cells, we compared the frequencies of targeted mutations only between parental 1C3-1 clone and tumor cell clones (Fig. 5B). The frequencies of some targeted mutations changed significantly among these tumor cells (Fig. 5B; Additional file 1: Table S6). For instance, 1 bp deletion (Del1|0) of Rb1 was dominant with the frequencies at over 50% in SG1-1, SG1-2, SG2-5, and SG2-6 but infrequent at less than 5% in SG4-1, SG4-3, SG5-1, and SG5-2 (Fig. 5C). Differently, the parental subclone 1C3-1 harbored this Rb1 Del1|0 mutation at 40% (Fig. 5C). Furthermore, while the Del5|2 mutation of Lkb1 changed little with the frequency at about 40% among all tumor cells tested, the Lkb1 Ins1G mutation oscillated significantly among these tumor cells, with the frequency at about 40% in SG1-1, SG1-2, SG2-5, and SG2-6 as well as 1C3-1 but nearly undetectable in SG4-1, SG4-3, SG5-1, and SG5-2 (Fig. 5D). In addition, two Col1a1 mutations, i.e., Del1|0 and Ins1T, oscillated in an opposite direction (Fig. 5E). While the Col1a1 Del1|0 mutation dominant in 1C3-1, SG1-1, SG1-2, SG2-5, and SG2-6 was negligible in SG4-1, SG4-3, SG5-1, and SG5-2, the Col1a1 Ins1T mutation infrequent in 1C3-1, SG1-1, SG1-2, SG2-5, and SG2-6 occurred frequently in SG4-1, SG4-3, SG5-1, and SG5-2 (Fig. 5E). Similarly, the Del1|0 and Ins1A mutations of Rasa1 started with the frequencies at around 50% in parental 1C3-1 clone and then oscillated in a reverse pattern among single-cell clones from tumor grafts (Fig. 5F). These results indicate that targeted mutations in tumor cells derived from CRISPR/Cas9-induced liver cancer in mice are also unstable in subcutaneous grafts derived from tumor cells with the frequencies of some targeted mutations oscillating in vivo.
Fig. 5
Oscillation of target site mutations during subcutaneous tumor cell grafts of 1C3-1. A Schematic of experimental outline. After subcutaneous implantation of the single-cell subclone 1C3-1 into SCID mice, tumors were formed and four tumor tissues (SG1, SG2, SG4, and SG5) from different mice collected at 2 weeks post inoculation. Two single-cell clones were derived from each of these four tumor tissues as indicated. Target site mutations were determined by targeted PCR amplicon deep sequencing of gDNA extracted from cells. SG, subcutaneous graft. B Changes in target site mutation profiles of single-cell clones from subcutaneous tumor cell grafts as indicated. Gene targets and their mutation types are shown on top and at bottom, respectively. The numbers flanking Del or Ins indicated at bottom are the numbers of deleted or inserted nucleotides. √Frequency representing square root of mutation frequencies is indicated from the highest 1 in dark green to the lowest 0 in white. The top line chart and the boxplot under show the number of single-cell clones with a specific mutation type, i.e., number of clones, and the percentage distribution of a specific mutation type in all single-cell clones, i.e., proportion (%), respectively. C–F Representative target mutation oscillation of Rb1 (C), Lkb1 (D), Col1a1 (E), and Rasa1 (F) across single-cell clones derived from subcutaneous tumor cell grafts
Increased genomic instability in CRISPR/Cas9-induced mouse liver cancer cellsAs mentioned previously [52], spontaneous mutations at a given site normally occur at an extremely low rate and are impossible to cause significant alterations in the types and frequencies of targeted mutations detected in the study. Continuing targeted editing by Cas9-sgRNA was also excluded as a causal factor because neither stable expression nor the editing activity of Cas9-sgRNA was detected in these single-cell clones. As cancer with genomic instability has an increased tendency for constant genomic alteration, we wondered whether these primary liver tumor cells are associated with strong genomic instability. We first examined spontaneous γ-H2AX and 53BP1 foci formation in 10 primary tumor cell lines derived from CRISPR/Cas9-induced mouse liver cancer, the control cell line NIH3T3, and the mouse liver cancer cell line Hepa1-6 (Fig. 6A). All of these liver cancer cell lines except the 1C3-2 clone showed a higher level of spontaneous γH2AX focus formation than the control NIH3T3 cells (Fig. 6A–B), indicating strong induction of spontaneous DNA DSBs and activation of DNA damage response in these cancer cells. It was however unclear why 53BP1 focus formation was much less frequent than γH2AX focus formation in nearly all of these cell lines as both γH2AX and 53BP1 foci indicated the site of DSBs (Fig. 6A–B).
Fig. 6
Genomic instability in single-cell clones derived from CRISPR/Cas9-induced mouse liver tumors. A Microscopic images of spontaneous γH2AX and 53BP1 foci in primary mouse liver cancer cells. B Percentage of cells with > 5 γH2AX foci or 53BP1 foci. Columns indicate the mean ± S.E.M from three independent experiments with statistical significance detected by One-way ANOVA with Dunnett’s multiple comparison test (vs. NIH-3T3). *P < 0.05 and ***P < 0.001. C Microscopic images of micronuclei (MN) in primary mouse liver cancer cells. D Percentage of cells with micronuclei. Columns indicate the mean ± S.E.M from three independent experiments with statistical significance detected by One-way ANOVA with Dunnett’s multiple comparison test (vs. NIH-3T3). **P < 0.01 and ***P < 0.001. E Representative images of chromosomal aberrations in metaphase spread of primary mouse liver cancer cell lines. Red arrows and black arrows indicate chromosomal fragments and biarmed chromosomes, respectively. F Summary of metaphase spread analysis. In each indicated cell line, the number of metaphases, number of chromosomes per metaphase, and percentages of metaphases with biarmed chromosomes and with chromosomal fragments are calculated and shown
We also analyzed these CRISPR/Cas9-induced primary liver tumor cells for micronucleus formation, which is frequently involved in chromosomal aberrations and genomic instability in cancer [53]. Micronucleus formation was readily detected in these tumor cells (Fig. 6C). The percentages of micronucleated cells were significantly higher in tumor cells than in the control NIH3T3 cells (Fig. 6D). In particular, while about 1% of NIH3T3 cells contained micronuclei, over 10% of 1C3 or 1C3-1 cells were micronucleated (Fig. 6D). Metaphase spread analysis further identified significant chromosomal aberrations in primary liver tumor cells (Fig. 6E). The number of chromosomes in primary liver tumor cells varied in average from 57.4 in 1C3 to 105.7 in 6C7-2 and was much more than 40 in a diploid mouse cell (Fig. 6E–F). This indicates that these tumor cells are hyperploid, at least in part contributing to more than two allelic variations at a target site of TSGs in single-cell clones. Biarmed chromosomes appeared in all Hepa1-6 cells as reported previously [54], but only existed in 2.0–6.0% of the primary liver tumor cell lines 1C3, all 1C3 subclones, and 6C7-5 as well as in 8.9% of NIH-3T3 (Fig. 6E–F). In contrast, we did not detect any biarmed chromosomes in 6C7 or its subclones 6C7-2, 6C7-3, and 6C7-4. The difference in the generation of biarmed chromosomes between 6C7-5 and its parental clone 6C7 or other 6C7 subclones indicates potential variations in genomic instability in these cells (Fig. 6E–F). In addition, primary liver tumor cells exhibited more frequent chromosomal fragments than NIH-3T3 and Hepa1-6 (Fig. 6E–F). As frequent micronucleus formation, chromosomal aberrations, and chromosomal fragments in primary tumor cells manifested the genomic instability of CRISPR/Cas9-induced mouse liver tumors, these findings indicated a possible connection between the genomic instability and the oscillation in the frequencies of targeted TSG mutations in tumor cells. Variations in genomic instability among parental clones (i.e., 1C3 and 6C7) and their respective subclones again implied that an intrinsic genetic force drive the oscillation of target site mutations in these single-cell clones.
Detectable eccDNA harboring targeted site mutationsDue to unstable nature, linear chromosomal fragments observed in our study could drive significant alterations in the frequencies of targeted mutations in tumor cells derived from CRISPR/Cas9-induced mouse liver cancer. However, it was technically difficult to separate linear chromosomal fragments from intact chromosomes for sequencing. Thus, we were unable to determine the extent to which linear chromosomal fragments contributed to the oscillation in the frequencies of targeted mutations in tumor cells. Like linear chromosomal fragments, eccDNA could be formed in CRISPR/Cas9-induced mouse liver cancer; however, as eccDNA could be separated from gDNA and identified by Circle-Seq [45], it was possible to determine the contribution of eccDNAs in the oscillation of the targeted TSG mutation frequencies in tumor cells.
We thus isolated eccDNAs from the 1C3-1 clone and the 6C7 clone along with the control NIH3T3 cells, enriched eccDNAs by removal of linear DNA with Plasmid-Safe ATP-Dependent DNase and removal of mitochondrial DNA with PacI and RCA of eccDNAs with Phi29 DNA polymerase, and performed Circle-Seq followed by Circle-MAP (Additional file 2: Fig. S7A). Removal of linear DNA by Plasmid-Safe ATP-Dependent DNase and mitochondrial DNA by PacI from eccDNAs was confirmed by targeted PCR amplification of the nuclear gene Actb and Cox5b and the mitochondrial gene mt-Co1 (Additional file 2: Fig. S7B-D). Circle-MAP revealed that the numbers of eccDNAs were respectively 12,637 in 1C3-1 and 12,611 in 6C7, greater than 7282 in NIH3T3 (Fig. 7A; Additional file 1: Table S7). The length distribution of eccDNAs was similar in all three cell lines, and the majority of these eccDNAs were small in size (Fig. 7A). For example, the length of eccDNA ranged from 150 to 9993 bp with the median length of 344 bp in the 1C3-1 clone, from 150 to 9962 bp with the median length of 466 bp in the 6C7 clone, and from 150 to 9987 bp with the median length of 333 bp in the control NIH3T3 cells (Fig. 7A); 83%, 68%, and 86% were less than 1000 bp and 91%, 82%, and 93% less than 2000 bp in 1C3-1, 6C7, and NIH3T3, respectively (Fig. 7A; Additional file 1: Table S7). Additionally, eccDNAs were mostly distributed in distal intergenic region and introns; however, the distribution of eccDNAs in genic regions such as promoters, 5′-UTRs, 3′-UTRs, and exons was more frequent in 1C3-1 and 6C7 cells than in NIH3T3 cells (i.e., 15.78% in 1C3-1 and 17.81% in 6C7 vs. 13.78% in NIH3T3).
Fig. 7
Identification and mapping of eccDNAs derived from the single-cell clone 1C3-1 and 6C7 by Circle-Seq. A EccDNA size distribution and mapping of eccDNAs to genic and intergenic regions in 1C3-1 (top) and 6C7 (middle) as well as in the control NHH-3T3 cell line (bottom). Total number of eccDNAs are indicated in parentheses next to the name of the cell line. Pie charts depict the distribution and percentage of eccDNAs mapped to different genic and intergenic regions in the three cell lines. Percentages of eccDNAs are also shown for genic regions that include promoters, exons, 5′-UTR and 3′-UTR. B Distribution of eccDNAs neighboring TSG target sites in various distances indicated by log10 values from distal and proximal junction point to TSG target sites in NIH-3T3, 1C3-1, and 6C7 cell lines as indicated. EccDNAs that are close to TSG target sites are also denoted by arrows and names. X and Y in the schematic (top) represent the distance from the distal and proximal junction point of eccDNAs on the chromosome to the breakpoint of TSG target site by CRISPR/Cas9, respectively. C Schematic for circularization of Apob-2119 eccDNA from chromosome 12 (Chr 12). Likely due to cleavage of the Apob target site by CRISPR/Cas9, the left end was processed and circularized via neighboring microhomology (MH) in the same chromosome to form Apob-2119 eccDNA. The four primers F1, R1, F2, and R2 indicated were designed to form one pair of inward primers F2/R2 and two pairs of outward primers R1/F1 and R2/F1 for validation of Apob-2119 eccDNA. The length of the eccDNA and the position of Apob target site by CRISPR/Cas9 are shown with the distal point of the eccDNA set at 0 bp. D Sequences of Apob-2119 eccDNA and its junctions as shown are determined by PCR with indicated primer pairs followed by Sanger sequencing. Circularization junction is also indicated
We also mapped eccDNAs to 24 chromosomes and determined the chromosomal regions from which each eccDNA was likely originated (Additional file 1: Table S7; Additional file 2: Fig. S8A). After selecting eccDNAs that were matched to target TSG regions as well as the Setd5 control (Additional file 1: Table S7; Additional file 2: Fig. S8B-D), we analyzed the distance from the breakpoints at Cas9-sgRNA target sites of TSGs and Setd5 to the sites that were corresponding to the distal and proximal junction of eccDNAs. The distance was generally shorter in the 1C3-1 clone and the 6C7 clone than in the control NIH3T3 cells (Fig. 7B; Additional file 1: Table S7). Among eccDNAs near target TSGs, the 435-bp Apob-2119 eccDNA near Apob was the closest eccDNA to a target TSG in 1C3-1 cells (Fig. 7B; Additional file
留言 (0)