
記住我
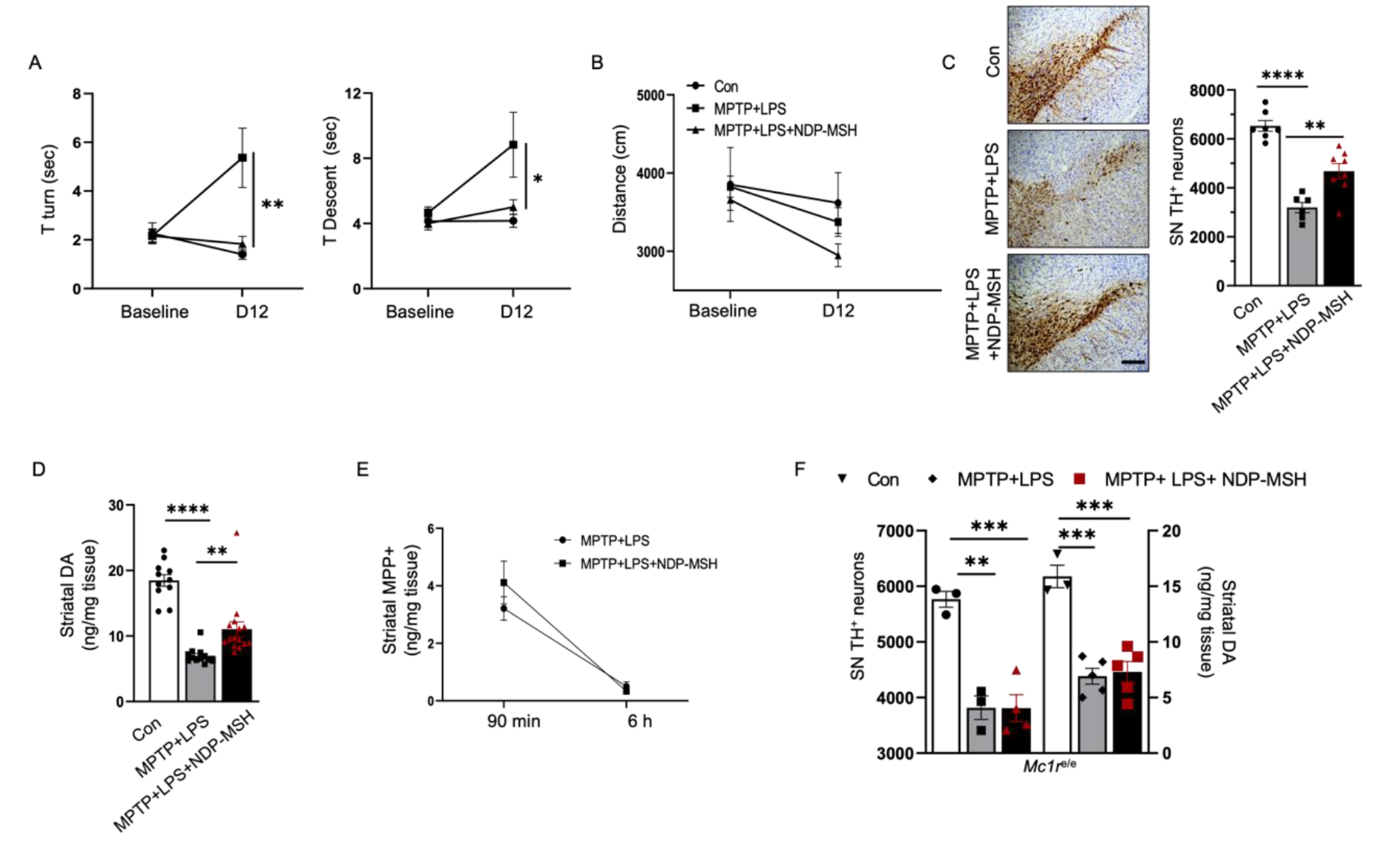





To assess the pharmacological impact of Re on photooxidative stress, the level of ROS was examined in the retina in situ 3 h after the experimental light exposure and the indicated treatments, which included Re treatment administered at 5, 25 and 100 mg/kg. As shown in the Fig. 1a and b, the ROS signal was barely detected in the outer nuclear layer (ONL) in the retinas from the normal controls; however, the level of ROS was evidently increased in the ONL in the vehicle-treated light-exposed retinas. On the other hand, reductions in the ROS signal in the ONL were readily detected in the light-exposed mice treated with Re at 25 and 100 mg/kg. Given that comparable levels of ROS signal in the ONL were noted in the normal controls and the light-exposed mice treated with 100 mg/kg Re, Re treatment was repeated at 100 mg/kg, followed by examination of the retinal ROS production 1 d after the experimental light exposure. As shown in Fig. 1c and d, the pronounced increase in the ROS signal was consistently observed in the ONL in the light-exposed vehicle-treated retinas compared to the normal controls. In sharp contrast, much less ROS signal was detected in the ONL in the light-exposed Re-treated retinas. Furthermore, given that the mechanisms of the anti-oxidant properties of Re implicate alleviating lipid peroxidation (Verstraeten et al. 2020) and lipid peroxidation is a major consequence of oxidative stress in photoreceptors, IHC was further performed to examine the immunopositivity of 4HNE, a marker for lipid peroxidation, in the retina. As shown in Fig. 1e and f, increased immunopositivity of 4HNE was readily detected in the ONL and retinal pigment epithelium (RPE) in the light-exposed vehicle-treated retinas compared to the normal controls. In contrast, significantly decreased 4HNE immunopositivity was noted in the light-exposed Re-treated retinas compared to the light-exposed vehicle-treated retinas. These results collectively indicate that Re is effective at attenuating photooxidative stress in the retina.
Fig. 1
Re-treatment attenuates ROS production and lipid peroxidation in the light-exposed retinas. (a) Retinal ROS production (in red) 3 h after light exposure. DAPI positivity (in blue) highlighted nuclei. NL, the vehicle-treated mice unexposed to the experimental light exposure; LD, the light-exposed vehicle-treated mice; Re_L, the light-exposed mice treated with 5 mg/kg Re; Re_M, the light-exposed mice treated with 25 mg/kg Re; Re_H, the light-exposed mice treated with 100 mg/kg Re. Scale bar, 50 μm. (b) Quantification of ROS signal in the ONL. Relative fold change was plotted against NL. (c) Retinal ROS production (in red) 1d after light exposure. Re, the light-exposed mice treated with 100 mg/kg Re. Scale bar, 50 μm. (d) Quantification of ROS signal in the ONL. Relative fold change was plotted against NL. (e) IHC of 4HNE immunopositivity in the retina 1d after the light exposure. Scale bar, 50 μm. (f) Quantification of the immunopositivity of 4HNE in the ONL. Relative fold change was plotted against NL. Data were expressed as mean ± SEM (n = 6 per group). ## Compared to NL, P < 0.01; ### compared to NL, P < 0.001; * compared to LD, P < 0.05; *** compared to LD, P < 0.001; Ɨ compared to NL, p < 0.05; ƗƗƗ compared to NL, p < 0.001. ns, not significant. INL, inner nuclear layer; ONL, outer nuclear layer; RPE, retinal pigment epithelium
Re Protects Against Photooxidative Stress-Mediated Impairment of the Photoreceptor Morphological IntegrityAs shown above, Re treatment attenuates light-induced photooxidative stress in the retina. Given that photooxidative stress is the key mechanism leading to photoreceptor degeneration, the potential protective effects of Re against photooxidative stress-mediated loss of photoreceptors were further assessed using full-retinal cross-sectional OCT scans after the indicated treatments. The experimental light exposure resulted in selective diminishment of the photoreceptor IS/OS and ONL in both superior and inferior retinas without causing overt morphological changes in the inner retina. In contrast, no significant changes in the photoreceptor morphological integrity were observed in the light-exposed Re-treated mice (Fig. 2a). Consistently, the ONL and IS/OS but not the inner nuclear layer (INL) was much thinner in the superior and inferior retinas in the light-exposed vehicle-treated retina compared to the normal controls, whereas the thickness of ONL and IS/OS was increased in the inferior and superior retinas in the light-exposed mice Re-treated mice (Fig. 2b). Furthermore, the protection of Re against photooxidative stress-mediated photoreceptor degeneration was confirmed following HE examination of the retinal gross morphology. Obliteration of the photoreceptor OS, IS and ONL was observed in the light-exposed vehicle-treated retinas; however, the morphological features of the photoreceptor OS, IS and ONL were to a large extent preserved in the light-exposed Re-treated mice (Supplemental Fig. 1a). Consistently, the number of photoreceptor nuclei was significantly decreased in the light-exposed vehicle-treated retinas. In sharp contrast, the number of photoreceptor nuclei was remarkably increased in the light-exposed Re-treated mice (Supplemental Fig. 1b). Therefore, the results from the OCT imaging and HE examination collectively demonstrate that Re protects against photooxidative stress-triggered photoreceptor morphological degeneration.
Fig. 2
Re-treatment attenuates morphological impairment of photoreceptors in the light-exposed retinas. (a) OCT scans of the retina. NL, the vehicle-treated mice unexposed to the experimental light exposure; LD, the light-exposed vehicle-treated mice; Re, the light-exposed mice treated with 100 mg/kg Re. (b) The thickness of the ONL, IS/OS and INL at 500 μm off the ONH in the superior and inferior retina. White asterisk, impaired ONL. Closed triangle, IS/OS. Scale bar, 100 μm. Data were expressed as mean ± SEM (n = 6 per group). ### Compared to NL, P < 0.001; *** compared to LD, P < 0.001; ns, not significant. GCL, ganglion cell layer; INL, inner nuclear layer; IPL, inner plexiform layer; IS, inner segment; ONH, optic nerve head; ONL, outer nuclear layer; OPL, outer plexiform layer; OS, outer segment; RPE, retinal pigment epithelium
Re Maintains the Retinal Function Under Photooxidative Stress ConditionsNext, ERG was performed to further evaluate the pharmacological impact of Re on the retinal function under photooxidative stress conditions. In the retinas from the normal controls, the flash of light induced a typical biphasic waveform consisting of the first negative wave component, a-wave, and the following positive wave component, b-wave. In distinct contrast, this biphasic ERG waveform was evidently flattened in the light-exposed vehicle-treated retinas. However, the ERG responses from the light-exposed Re-treated retinas yielded biphasic ERG waveform patterns similar to the normal controls (Fig. 3a). Meanwhile, light stimuli induced intensity-dependent increases in the scotopic a-wave (Fig. 3b) and b-wave amplitudes (Fig. 3c) in the normal controls. Consistently, light intensity-dependent increases in the a-wave and b-wave amplitudes were markedly reduced in the light-exposed vehicle-treated retinas compared to the normal controls. The a-wave and b-wave amplitudes in the light-exposed Re-treated retinas were significantly increased compared to the light-exposed vehicle-treated retinas (Fig. 3b and c). Thus, the results from the ERG recording demonstrate that Re effectively maintains the retinal function under photooxidative stress conditions.
Fig. 3
Re maintains the retinal function and preserves the expression of rhodopsin in the light-exposed retinas. (a) Representative electroretinograms from scotopic ERG recordings. (b) Amplitudes of a-wave. (c) Amplitudes of b-wave. (d) IHC of rhodopsin (in red) and DAPI positive nuclei (in blue). White asterisk, the diminished ONL. White arrows, mislocalized rhodopsin. Scale bar, 50 μm. Data were expressed as mean ± SEM (n = 6 per group). ### Compared to NL, P < 0.001; *** compared to LD, P < 0.001. INL, inner nuclear layer; IS, inner segment; ONL, outer nuclear layer; OS, outer segment
Well-maintained ERG responses prompted us to further probe the histopathological mechanisms of Re-conferred retinal functional protection under photooxidative stress conditions. As photoreceptors contribute to the a-wave response in ERG (Breton et al. 1994), IHC was performed to assess the impact of Re treatment on the morphology of rod and cone photoreceptors in situ. First, we examined the expression of signature proteins expressed in the photoreceptor OS, including rhodopsin, an essential functional protein of rod photoreceptors, S-opsin, a marker for the short-wavelength-sensitive cone photoreceptors and M-opsin, a marker for the middle- and long-wavelength-sensitive cone photoreceptors. As shown in Fig. 3d, abundant expression of rhodopsin was readily detected in the OS in the normal controls. In sharp contrast, the OS-specific expression pattern of rhodopsin was overtly disrupted in the light-exposed vehicle-treated retinas 6 h and 1 d after the light exposure. By 3 d post the experimental light exposure, the expression of rhodopsin was remarkably diminished in the light-exposed vehicle-treated retinas. By 7 d after the light exposure, the expression of rhodopsin was barely detected in the light-exposed vehicle-treated retinas. On the contrary, Re treatment maintained the normal expression pattern of rhodopsin in the light-exposed retinas. Similar observations were made with respect to the expression pattern of S-opsin (Supplemental Fig. 2a) and M-opsin (Supplemental Fig. 2b). The results here demonstrate that Re treatment leads to well-maintained photoreceptor OS morphology under photooxidative stress conditions.
ERG b-wave reflects the function of bipolar cells (Stockton and Slaughter 1989). Our previous studies have demonstrated that during the course of photooxidative stress-mediated photoreceptor degeneration, the second-order retinal neurons bipolar cells manifest morphological impairment, which may in part contribute to the functional deterioration of the retinas (Chen et al. 2016). Thus, the effect of Re treatment on the morphology of the bipolar cells was also examined to further characterize the retinal protective effects of Re. The bipolar cells can be visualized by the expression of PKCα in the retina. IHC examination revealed that the dendrites of bipolar cells were markedly impaired (Supplemental Fig. 3a) and the immunopositivity of PKCα in the outer plexiform layer (OPL) was significantly reduced (Supplemental Fig. 3b) in the light-exposed vehicle-treated retinas compared to the normal controls. In contrast, increased immunopositivity of PKCα in the OPL outlined well-preserved bipolar cell dendritic terminals in the light-exposed Re-treated retinas (Supplemental Fig. 3a and b). These results indicate that Re treatment protects against photoreceptor degeneration-associated impairment of the second-order bipolar cells under photooxidative stress conditions.
Re Treatment Counteracts the Deleterious Impact of Photooxidative Stress on the Retinal Gene ExpressionNext, the retinas were collected at 1 d post experimental light exposure and subjected to RNA-seq analyses to further understand the mechanisms associated with the retinal protective effects of Re against photooxidative stress. Principal component analysis (PCA) provided an unbiased overview that the retinal gene expression profiles of the normal controls and that of the light-exposed vehicle-treated mice were overtly separated. However, the retinal gene expression profiles from the light-exposed Re-treated mice clustered closer to that from the normal controls (Fig. 4a). Consistently, hierarchical clustering analysis of the differentially expressed genes (DEGs) revealed that unlike the gene expression profiles from the light-exposed vehicle-treated retinas, the retinal gene expression profiles in the light-exposed Re-treated mice were similar to the normal controls (Fig. 4b). Additionally, gene set enrichment analysis (GSEA) of KEGG and GO revealed that experimental light exposure resulted in significant upregulation in the pathways such as neuroinflammatory response, microglial cell activation and cellular response to oxidative stress in the retina. On the other hand, pathways including chromatin silencing, retina homeostasis, visual perception, photoreceptor inner segment, photoreceptor cell cilium, photoreceptor outer segment and phototransduction were significantly downregulated in the light-exposed vehicle-treated retinas (Fig. 4c). However, compared to the light-exposed vehicle-treated retinas, the light-exposed Re-treated retinas were characterized by significantly downregulated pathways of neuroinflammatory response, microglial cell activation and cellular response to oxidative stress as well as upregulated pathways of chromatin silencing, retina homeostasis, visual perception, photoreceptor inner segment, photoreceptor cell cilium, photoreceptor outer segment and phototransduction (Fig. 4d). These results provide molecular evidence that further corroborates the retinal protective effects of Re against photooxidative stress-mediated photoreceptor degeneration.
Fig. 4
Re-treatment counteracts light-induced alterations of the gene expression profiles in the retina. (a) PCA of all data sets from NL (gray), LD (green) and Re (blue) (n = 3 per group). (b) Hierarchical clustering analysis of the DEGs from NL, LD and Re. Scale bar, the Z score indicative of upregulation (red) and downregulation (blue) of gene expression. (c) GSEA based on KEGG or GO gene sets (LD vs. NL). (d) GSEA based on KEGG or GO gene sets (Re vs. LD). FDR, false discovery rate; NES, normalized enrichment score; pval, p-value
A further analysis of the pathways associated with oxidative stress unveiled that gene sets such as reactive oxygen species metabolic process, superoxide metabolic process, hydrogen peroxide metabolic process, reactive nitrogen species metabolic process, cell death in response to oxidative stress and respiratory burst were significantly upregulated in the light-exposed vehicle-treated retinas (Fig. 5a). In contrast, these gene sets were noted to be significantly downregulated in the light-exposed Re-treated retinas compared to the light-exposed vehicle-treated retinas (Fig. 5b). Taken together, the results from the transcriptomic analyses provide a non-biased overview of the molecular pathways associated with Re-conferred retinal protection under photooxidative stress conditions.
Fig. 5
Re treatment counteracts light-induced alterations of the gene sets associated with oxidative stress in the retina. (a) Positively enriched oxidative stress-related gene sets in the light-exposed vehicle-treated retinas as revealed by GSEA. (b) Negatively enriched oxidative stress-related gene sets in the light-exposed Re-treated retinas as revealed by GSEA. FDR, false discovery rate; NES, normalized enrichment score; pval, p-value
Re Treatment Counters Photooxidative Stress-Triggered Downregulation of Genes Implicated in PhototransductionPhotoreceptors are specialized retinal neurons equipped with the function to convert light into electrical signals, a process defined as phototransduction. Phototransduction initiates visual processes and thereby is indispensable for vision formation (Mannu 2014). As shown above, Re treatment counteracted photooxidative stress-triggered downregulation of phototransduction gene set in the retina (Fig. 4d), corroborating the functional preservation conferred by Re treatment (Fig. 3a-c). Given the critical role of phototransduction in the retinal function, real-time qPCR was further performed to validate the impact of Re treatment on the retinal expression of phototransduction gene signatures revealed by RNA-seq analyses, including rod cyclic GMP-gated cation channel subunit encoding gene Cnga1, rod-specific Gα transducin subunit encoding gene Gnat1, guanylate cyclase activator protein GCAP1 encoding gene Guca1b, M-opsin encoding gene Opn1mw, S-opsin encoding gene Opn1sw, phosphodiesterase 6 encoding gene Pde6b, rhodopsin encoding gene Rho, and Slc24a1, which encodes a member of the potassium-dependent sodium/calcium exchanger protein family. Consistent with the results obtained from the RNA-seq analyses (Fig. 6a), real-time qPCR revealed significantly decreased retinal expression of Cgna1, Gnat1, Guca1b, Opn1mw, Opn1sw, Pde6b, Rho, and Slc24a1 in the light-exposed vehicle-treated mice compared to the normal controls. In sharp contrast, significantly increased retinal expression of Cgna1, Gnat1, Guca1b, Opn1mw, Opn1sw, Pde6b, Rho, and Slc24a1 was noted in the light-exposed Re-treated mice compared to the light-exposed vehicle-treated mice (Fig. 6b). These results confirm that Re treatment counteracts photooxidative stress-triggered downregulation of genes implicated in phototransduction.
Fig. 6
Re maintains the retinal expression of genes essential for phototransduction in the light-exposed retinas. (a) The retinal expression of phototransduction genes revealed by RNA-seq (n = 3 per group). (b) Real-time qPCR validation of the retina expression of phototransduction genes (n = 6 per group). Relative fold change was plotted against NL. Data were expressed as mean ± SEM. # Compared to NL, P < 0.05; ## compared to NL, P < 0.01; ### compared to NL, P < 0.001; ** compared to LD, P < 0.01; *** compared to that from LD, P < 0.001
Re Attenuates Photoreceptor Degeneration-Associated Inflammation and Microglial Activation in the RetinaBased on the results from the RNA-seq analyses, neuroinflammatory response and microglial cell activation are the most upregulated pathways in the retina in response to the damaging light exposure (Fig. 4c). Re treatment resulted in a significant downregulation of gene sets of neuroinflammatory response and microglial cell activation in the retina (Fig. 4d). Mounting evidence has supported the pathophysiological significance of microglial activation and inflammation in the progression of photoreceptor degeneration (Rashid et al. 2019). Therefore, real-time qPCR was performed to validate the expression of neuroinflammatory genes including Ccl2, Ccl3, Ccl4, Il1b, Tlr4 and Tnf as well as Axl, Cd68, Clec7a and Tspo, genes closely associated with microglial activation as revealed by the RNA-seq analyses (Fig. 7a). As shown in Fig. 7b, significantly elevated expression of Ccl2, Ccl3, Ccl4, Il1b, Tlr4, Tnf, Axl, Cd68, Clec7a and Tspo was observed in the light-exposed vehicle-treated retinas compared to the normal controls. In contrast, the retinal expression of these genes was significantly decreased in the light-exposed Re-treated mice compared to their light-exposed vehicle-treated counterparts. In addition, the immunopositivity of Iba1 and CD68 was also examined by IHC to visualize the changes of microglia in situ. The Iba1 positive cells were ectopically present in the ONL and the subretinal space in the light-exposed vehicle-treated mice 1 d and 3 d after the experimental light exposure. The ectopic localization of Iba1 positive cells in the ONL and the subretinal space was significantly attenuated in the light-exposed Re-treated retinas (Fig. 7c and d). Similar observations were made for the CD68 immunopositivity in the ONL and the subretinal space (Fig. 7e and f). Taken together, these results indicate that Re treatment suppresses microglial activation and neuroinflammatory responses in the light-exposed retinas.
Fig. 7
Re treatment inhibits neuroinflammation and microglial activation in the light-exposed retinas. (a) The retinal expression of the genes associated with neuroinflammation and microglial cell activation revealed by RNA-seq (n = 3 per group). Relative fold change was plotted against NL. (b) Real-time qPCR validation of the retinal expression of the genes associated with neuroinflammation and microglial cell activation (n = 6 per group). Relative fold change was plotted against NL. (c) IHC examination of Iba1 (in red) and DAPI positive nuclei (in blue). Scale bar, 50 μm. (d) Quantification of the Iba1 immunopositivity in the ONL and subretinal space. Relative fold change in the Iba1 immunopositivity was plotted against LD 6 h. (e) IHC examination of CD68 (in red) and DAPI positive nuclei (in blue). Scale bar, 50 μm. (f) Quantification of the CD68 immunopositivity in the ONL and subretinal space. Relative fold change in the CD68 immunopositivity was plotted against LD 6 h. Data were expressed as mean ± SEM. # Compared to NL, P < 0.05; ## compared to NL, P < 0.01; ### compared to NL, P < 0.001; * compared to LD, P < 0.05; ** compared to LD, P < 0.01; *** compared to LD, P < 0.001; ns, not significant
Re Preserves Müller Cell Homeostasis Under Photooxidative Stress ConditionsIn addition to the changes manifested by microglia during the course of photoreceptor degeneration, gliotic remodeling of the principal macroglia in the retina, müller cells, is encountered in nearly all types of retinal diseases including photoreceptor degeneration (Bringmann et al. 2006). Aside from causing functional impairment of the müller cells, reactive gliosis of müller cells may lead to the formation of the glial scar that in part contribute to deterioration of the retinal functional and structural integrity (Bringmann and Reichenbach 2001; Bringmann et al. 2006). Thus, the impact of Re treatment on reactive gliosis of müller cells was further assessed. First, a search into the data from the above-mentioned RNA-seq analyses revealed significantly increased retinal expression of Gfap in the light-exposed vehicle-treated retinas compared to the normal controls. Meanwhile, the expression of Glul, which encodes glutamine synthetase, a key enzyme that carries out the primary function of müller cells in glial-neuronal transmitter recycling, was significantly decreased in the light-exposed retinas. The upregulation of Gfap and concurrent downregulation of Glul support gliotic müller cells are functionally compromised under photooxidative stress conditions. In contrast, much lower expression of Gfap and significantly increased expression of Glul were noted in the light-exposed Re-treated retinas compared to the light-exposed vehicle-treated retinas (Fig. 8a). Moreover, validation of the retinal expression Gfap and Glul by real-time qPCR yielded similar results (Fig. 8b). In addition, the retinal expression pattern of Gfap was examined by IHC to directly visualize müller cell reactive gliosis in situ. As shown in Fig. 8c, the expression of Gfap was restricted to the nerve fiber layer (NFL) in the vehicle-treated normal controls. In distinct contrast, Gfap immunopositivity was noted to extend aberrantly across all layers of the retina in the light-exposed vehicle-treated retinas. Meanwhile, the aberrant pattern of Gfap was readily detected 6 h after the experimental light exposure and persisted 7 d after the light exposure. In sharp contrast to the light-exposed vehicle-treated retinas, the expression pattern of Gfap remained restricted to the NFL in the light-exposed Re-treated retinas (Fig. 8c). Compared to the normal controls, significantly increased Gfap immunopositivity across the retina was observed 6 h, 1 d, 3 d and 7 d after the light exposure in the vehicle-treated mice. The Gfap immunopositivity was much lower in the light-exposed Re-treated retinas at all the time points examined (Fig. 8d). These results collectively indicate that Re treatment alleviates müller cell reactive gliosis under photooxidative stress conditions.
Fig. 8
Re treatment attenuates reactive gliosis of müller cells in the light-exposed retinas. (a) The retina expression of Gfap and Glul revealed by RNA-seq (n = 3 per group). Relative fold change was plotted against NL. (b) Real-time qPCR verification of the retinal expression of Gfap and Glul (n = 6 per group). Relative fold change was plotted against NL. (c) IHC examination of GFAP (in red) and DAPI positive nuclei (in blue). Scale bar, 50 μm. (d) Quantification of the GFAP immunopositivity in the retina by ImageJ. Relative fold change in the GFAP immunopositivity was plotted against NL. Data were expressed as mean ± SEM. # Compared to NL, P < 0.05; ## compared to NL, P < 0.01; ### compared to NL, P < 0.001; * compared to LD, P < 0.05; ** compared to LD, P < 0.01; *** compared to LD, P < 0.001
留言 (0)