
記住我





First, we sought to compare the pattern of lampbrush chromomeres with the distribution of A/B compartments along the chicken interphase chromosomes, which are characterized by multiple switches between A and B compartments. The results of comparing the distribution of A/B compartments in chicken embryonic fibroblasts (CEF) with the pattern of chromomeres along the lampbrush chromosomes GGA1–GGA7 are presented in Figs. 1, 2, 3, 4 and Additional file 1: Figure S1.
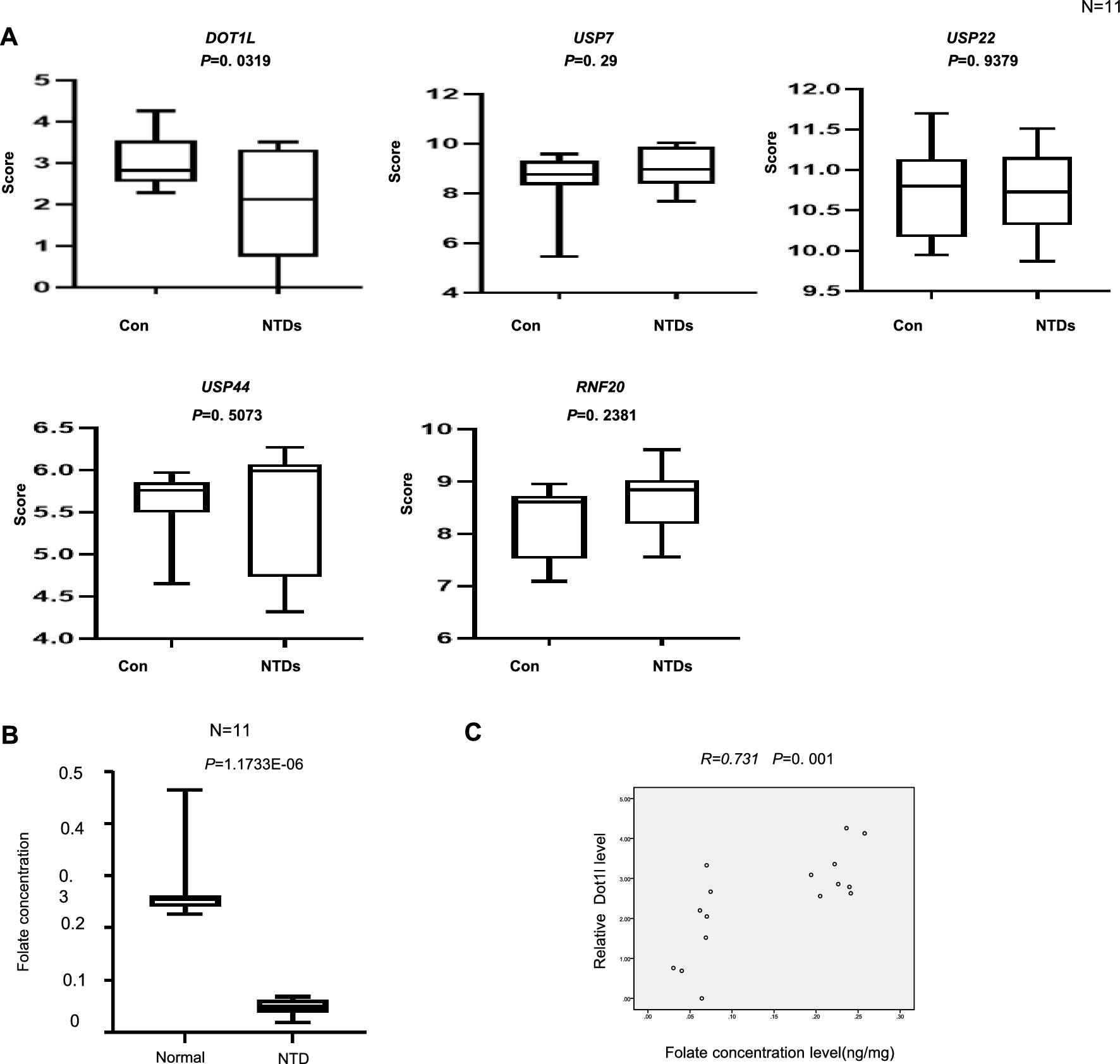
GGA1 lampbrush chromosome visually differs from the other macrochromosomes by its long length, the absence of any pronounced marker structures, with the exception of the terminal loops (telomere bow-like loops, TBLs) and one marker loop (proximal boundary of axial bar bearing no loops, PBL11) [52, 53]. In lampbrush chromosome 1 several clusters of dense compact chromomeres can be observed (Fig. 1a–c). Except for the pericentromeric cluster of chromomeres, all other clusters of prominent DAPI-positive chromomeres (clusters # 1, 2, 4, 5, as well as not numbered smaller regions) correspond to B compartments of different lengths in chicken embryonic fibroblasts.
Fig. 1
Alignment of interphase genome A/B compartments along chicken chromosome 1 with chromomeric pattern of the corresponding lampbrush chromosome. a—Distribution of A (red) and B (dark blue) compartments along the chicken chromosome 1 (GGA1) in embryonic fibroblasts viewed by Integrative Genomics Viewer (IGV) (according to [15]); b—cytological map of chicken lampbrush chromosome 1 depicting DAPI-staining pattern of chromomeres and relative contour length of lateral loops, black circles–dense chromomeres brightly stained with DAPI (according to [53, 54]). Dotted lines in a, b connect the genomic positions of the selected BAC-clones (Additional file 1: Table S1) with their positions on the cytological map; c—lampbrush chromosome 1 stained with DAPI, pixel intensities displayed with multicolored ImageJ look-up table, numbered lines in a and c indicate positions of chromomere clusters brightly stained with DAPI; d, e, f—DNA+RNA-FISH with BAC-clone based DNA-probes (Additional file 1: Table S2) covering the genomic regions 50–52 Mb (d), 70–71 Mb (e), and 185–186 Mb (f) on chicken lampbrush chromosome 1; dotted lines from c to d–f indicate chromosomal positions of the regions on microphotographs; dʹ–fʹ—the positions of the mapped BAC-clones relative to the somatic A/B compartments; dʹʹ–fʹʹ—schematic drawings of the FISH-mapping of the selected genomic regions on lampbrush chromatin domains; colors correspond to the colors of the labeled DNA-probes on the FISH images. Scale bar: c–20 μm, d–f—10 μm
Lampbrush chromosome GGA2 is characterized by the presence of the so-called ‘‘spaghetti’’ marker on the short arm and so-called lumpy loops (LLs) on the long arm [61]. DAPI-positive chromomeres are evenly distributed along the entire length of chromosome 2, with the large clusters in the centromeric region and on the long arm. The locus of ‘‘spaghetti marker’’ formation, the coordinates of which were established earlier [35], corresponds to the A compartment, whereas the cluster of prominent compact chromomeres, in which LLs form [65], corresponds to the B compartment in embryonic fibroblasts (Fig. 2a–c). The region of the extended cluster of dense compact chromomeres (cluster #5), the boundaries of which can be tentatively determined using previously mapped BAC clones [53, 54], corresponds to the large B compartment in embryonic fibroblasts (Fig. 2a–c).
Fig. 2
Alignment of interphase genome A/B compartments along chicken chromosome 2 with chromomeric pattern of the corresponding lampbrush chromosome. a—Distribution of A (red) and B (dark blue) compartments along the chicken chromosome 2 (GGA2) in embryonic fibroblasts viewed by Integrative Genomics Viewer (IGV) (according to [15]); b—cytological map of chicken lampbrush chromosome 2 depicting DAPI-staining pattern of chromomeres and relative contour length of lateral loops, black circles—dense chromomeres brightly stained with DAPI (according to [53, 54]). Dotted lines in a, b connect the genomic positions of the selected BAC-clones and chromosomal marker structures (Additional file 1: Table S1) with their positions on the cytological map; c—lampbrush chromosome 2 stained with DAPI, pixel intensities displayed with multicolored ImageJ look-up table, numbered lines in a and c indicate positions of chromomere clusters brightly stained with DAPI; d, e—DNA+RNA-FISH with BAC-clone based DNA-probes (Additional file 1: Table S2) covering the genomic regions 39–40 Mb (d), 128–135 Mb (e) on chicken lampbrush chromosome 2; dotted lines from c to d and e indicate chromosomal positions of the regions on microphotographs; dʹʹ, eʹ—the positions of the mapped BAC-clones relative to the somatic A/B compartments; d, eʹʹ—schematic drawings of the FISH-mapping of the selected genomic regions on lampbrush chromatin domains; colors correspond to the colors of the labeled DNA-probes on the FISH images. Scale bar: c—20 μm, d, e—10 μm.
The GGA3 lampbrush chromosome is well recognized due to the characteristic asymmetric length of the lateral loops, with a higher average loop length in the region of the long arm closer to the centromere, giving noticeable ‘‘fuzziness’’ in the proximal part of the long arm, whereas the distal part of the long arm is represented mainly by DAPI-positive chromomeres with short lateral loops [52, 53]. These features are well reflected in the distribution of epigenetic modifications: repressive chromatin markers H3K9me3 and 5-methyl-cytosine (5mC) predominate in the clusters of chromomeres in the second half of the long arm [38]. Extended clusters of large chromomeres in this region of lampbrush chromosome 3 generally correspond to the somatic B compartments (Additional file 1: Figure S1a–aʹʹ). For instance, the region around the LL marker of chromosome 3, which was mapped before [35], demonstrated the correspondence between 5 and 6 dense DAPI-positive chromomeres and homogenous B compartment in chicken embryonic fibroblasts.
Lampbrush chromosome GGA4, in turn, has well-defined terminal giant loops (TGLs) recently renamed GITERA (Giant Terminal RNP aggregates) [66], two clusters of DAPI-positive chromomeres, one in the short arm and one in the centromeric region, corresponding to B compartments in embryonic fibroblasts (Fig. 3a–c). Chicken lampbrush chromosome 4 is the most completely mapped, both by FISH with BAC clone-based probes [39, 53] and by microdissection of individual chromomeres followed by their sequencing [37]. In the proximal part of the long arm of the chromosome 4 is an extended cluster of small DAPI-positive chromomeres (cluster #3) that are depleted in acetylated histone H4 (H4Ac) and H3K9me3 [38]. This region of GGA4 corresponds to intermingled A/B compartments in chicken embryonic fibroblasts with the predominance of B compartment (Fig. 3a–c).
Fig. 3
Alignment of interphase genome A/B compartments along chicken chromosome 4 with chromomeric pattern of the corresponding lampbrush chromosome. a—Distribution of A (red) and B (dark blue) compartments along the chicken chromosome 4 (GGA4) in embryonic fibroblasts viewed by Integrative Genomics Viewer (IGV) (according to [15]); b—cytological map of chicken lampbrush chromosome 4 depicting DAPI-staining pattern of chromomeres and relative contour length of lateral loops, black circles—dense chromomeres brightly stained with DAPI (according to [53]). Dotted lines in a, b connect the genomic positions of the selected BAC-clones (Additional file 1: Table S1) with their positions on the cytological map; c—lampbrush chromosome 4 stained with DAPI, pixel intensities displayed with multicolored ImageJ look-up table, numbered lines in a and c indicate positions of chromomere clusters brightly stained with DAPI; d—DNA+RNA-FISH with BAC-clone based DNA-probes (Additional file 1: Table S2) covering the genomic region 34–37 Mb (d) on chicken lampbrush chromosome 4; dotted lines from c to d indicate chromosomal position of the region on microphotographs; dʹ—the positions of the mapped BAC-clones relative to the somatic A/B compartments; dʹ—schematic drawing of the FISH-mapping of the selected genomic region on lampbrush chromatin domains; colors correspond to the colors of the labeled DNA-probes on the FISH image. Scale bar: c—20 μm, d—10 μm.
GGA5 lampbrush chromosome is characterized by several clusters of larger and more globular chromomeres including a cluster without prominent lateral loops in the pericentromeric region [56]. Four of these clusters correlate with the B compartments present at chicken interphase genome (Additional file 1: Figure S1 b–bʹʹ).
GGA6 in a lampbrush configuration can be divided into two parts—with more compact larger chromomeres and relatively short lateral loops and with tiny chromomeres and longer transcription loops [53]. Prominent B compartment can be assigned to the first part of lampbrush chromosome 6 with higher chromatin compaction (cluster #1) (Additional file 1: Figure S1c–cʹʹ).
The GGA7 midichromosome, like the other midichromosomes, is characterized by small DAPI-negative chromomeres from which long lateral loops emerge. Based on the analysis of DAPI staining and counting of the average number of chromomeres, we drew an approximate cytological map of the chromomere pattern for GGA7 (Fig. 4a–c). Clusters of DAPI-positive chromomeres are found in the centromeric region, the position of which was previously established [57], and in the distal region of the long arm. B compartments are detected in the genomic region corresponding to these clusters, but further FISH mapping is required to verify their correspondence to the DAPI-positive chromomeres.
Fig. 4
Alignment of interphase genome A/B compartments along chicken chromosome 7 with chromomeric pattern of the corresponding lampbrush chromosome. a—Distribution of A (red) and B (dark blue) compartments along the chicken chromosome 7 (GGA7) in embryonic fibroblasts viewed by Integrative Genomics Viewer (IGV) (according to [15]); b—cytological map of chicken lampbrush chromosome 7 depicting DAPI-staining pattern of chromomeres and relative contour length of lateral loops, black circles–dense chromomeres brightly stained with DAPI. Dotted lines in a, b connect the genomic positions of the BAC-clones (Additional file 1: Table S1) with their positions on the cytological map; c—lampbrush chromosome 7 stained with DAPI, pixel intensities displayed with multicolored ImageJ look-up table, numbered line in a and c indicates position of chromomere cluster brightly stained with DAPI; d, dʹ–DNA+RNA-FISH with BAC-clone based DNA-probes (Additional file 1: Table S2) covering the genomic region 12–14 Mb on chicken lampbrush chromosome 7; dotted lines from c to d, d′ indicate chromosomal positions of the region on microphotographs; dʹʹ—schematic drawing of the FISH-mapping of the selected genomic region (d, dʹ) on lampbrush chromatin domains; colors correspond to the colors of the labeled DNA-probes on the FISH images. e—the positions of the mapped BAC-clones relative to the somatic A/B compartments. Scale bar: c—20 μm, d, dʹ—10 μm.
In summary, by comparing the pattern of lampbrush chromomeres with the distribution of A/B compartments (Figs. 1, 2, 3, 4a–c, Additional file 1: Figure S1), we have established that lampbrush chromosome segments characterized by higher chromatin compaction, more globular chromomeres, and lower transcriptional activity generally correlate with the B compartments present at interphase chromatin. At the same time, no obvious correspondence with either A or B compartment identity is found for smaller chromomeres with relatively long lateral loops.
FISH-mapping of the genomic regions belonging to A or B compartments on chicken lampbrush chromosomesNext, we decided to turn our attention to high resolution mapping of the genomic regions belonging to A compartments or regions transitional between A and B compartments on lampbrush chromosome preparations. Based on the available Hi-C data, we selected several regions of interest to obtain probes for FISH mapping of A/B compartments present in chicken interphase genome. For the most part (with a few exceptions), we focused on regions, where the compartment sign was uniform at the 1 to 3 Mb length scales, on the largest macrochromosomes (GGA1–4, GGA7) and one microchromosome (GGA14). Depending on the region, from 2 to 6 BAC clones from the CHORI-261 library were selected to cover the A or B compartment (Additional file 1: Table S2). The DNA probes derived from the BAC clones to the selected regions of interest were mapped on lampbrush chromosomes by 2D-FISH. FISH was performed according to the DNA/DNA + RNA in situ hybridization protocol so that DNA-probes also hybridized with the nascent transcripts on the lateral loops, allowing the hybridization signal to be observed along the entire length of the transcription loop.
Mapping genomic regions belonging to A compartmentsTo map the genomic regions belonging to A compartments we selected the following segments on four chromosomes: 50–52 Mb in GGA1, 39–40 Mb in GGA2, 12–14 Mb in GGA7, and 1–2 Mb in GGA14 (Additional file 1: Table S2).
Six BAC clones were selected for the 50–52 Mb region of chromosome 1 (Fig. 1dʹ). FISH mapping of the selected BAC clones on lampbrush chromosome 1 shows the presence of chromomere-loop complexes with long lateral loops in the region corresponding to A compartment (Fig. 1d–dʹʹ).
BAC clones CH261-163C1, CH261-63A12, and CH261-134B20 were selected to the 39–40 Mb region of chicken chromosome 2 (GGA2) (Fig. 2dʹ). All three probes hybridized with the RNP-matrix of the lateral loops (Fig. 2d–d′′).
Three BAC clones CH261-93F1, CH261-126G14, and CH261-38J23 selected to the 12–14 Mb region of chicken chromosome 7 (GGA7) were mapped into a single pair of long lateral loops with two transcriptional units separated by a chromatin nodule (Fig. 4d–dʹʹ). The presence of a chromatin nodule within the lateral loop can be explained by the fact that in the absence of transcribed genes, chromatin tends to persist in a compact, ‘‘closed’’ state, even without being anchored into a nearby chromomere. Chromatin nodules on the lateral loops of lampbrush chromosomes have been described previously at the morphological level as 5mC enriched transcript-free DNP regions with the characteristic nucleosomal organization [67] and by BAC-clone mapping as untranscribed DNA regions [39].
Two BAC clones, CH261-94D13, CH261-168C19 to a 1–2 Mb region of chicken chromosome 14 (GGA14) also belonging to A compartment hybridized with the RNP matrix of two long lateral loops (Fig. 5c–cʹʹ).
Fig. 5
Alignment of interphase genome A/B compartments along chicken chromosome 14 with chromomeric pattern of the corresponding lampbrush chromosome. a—Distribution of A (red) and B (dark blue) compartments along the chicken chromosome 14 (GGA14) in embryonic fibroblasts viewed by Integrative Genomics Viewer (IGV) (according to [15]); b—cytological map of chicken lampbrush chromosome 14 depicting DAPI-staining pattern of chromomeres and relative contour length of lateral loops, black circles—dense chromomeres brightly stained with DAPI (according to [54]). Dotted lines in a, b connect the genomic positions of the BAC-clones (Additional file 1: Table S1) with their positions on the cytological map; c, d—DNA + RNA-FISH with BAC-clone based DNA-probes (Additional file 1: Table S2) covering the genomic regions 1–2 Mb cʹ and 10–13 Mb dʹ on chicken lampbrush chromosome 14, dotted lines from b to c and d indicate positions of the regions on the cytological map; cʹ, dʹ—the positions of the mapped BAC-clones relative to the somatic A/B compartments; cʹʹ, dʹʹ—schematic drawing of the FISH-mapping of the selected genomic regions (c, d) on lampbrush chromatin domains; colors correspond to the colors of the labeled DNA-probes on the FISH images. Scale bar: 10 μm
Overall, all four genomic regions belonging to the A compartments in somatic cells were mapped to the relatively long lateral loops and adjacent rather small chromomeres in lampbrush chromosome chromatin.
Mapping genomic regions transitional between A and B compartmentsWe additionally selected BAC clones for A to B compartment transition regions in chicken embryonic fibroblasts for FISH-mapping on lampbrush chromosomes isolated from the oocytes. To this end, 70–71 Mb region in GGA1, 128–135 Mb region in GGA2, 34–37 Mb region in GGA4, and 10–13 Mb region in GGA14 were mapped (Additional file 1: Table S2).
The 70–71 Mb region of GGA1 is at the border between the chromosomal segment with very long lateral loops and the cluster of centromeric DAPI-positive chromomeres (the so-called ‘centromere bar’). This segment of the lampbrush chromosome 1 corresponds to the interphase chromatin region with multiple switches between A and B compartments (Fig. 1a–c). BAC clones CH261-162E14, CH261-33I15, and CH261-180H2 from this region hybridized with two neighboring chromomeres with pairs of short lateral loops (Fig. 1e–eʹʹ).
We also selected two differently labeled BAC clone sets to map an extended 128–135 Mb genomic segment of transition from the A to the B compartment on chicken chromosome 2 (GGA2) (Fig. 2eʹ). BAC clones CH261-98G7, CH261-135E13 picked up to the A compartment hybridized into chromomere-loop complexes with long lateral loops, indicating chromatin openness in this region. Next, BAC clones CH261-12O18, CH261-140E6, CH261-177C13, CH261-97C18, and CH261-54I9, matched to the B compartment, hybridized into chromomeres (Fig. 2e–eʹʹ). One of these BAC clones also hybridized with a very short lateral loop, which may be related to the transcription of a certain gene. This hybridization pattern fully confirms our observation about the preferential correspondence of A compartments to smaller chromomeres with longer lateral loops and B compartments—to larger chromomeres with shorter lateral loops.
Two other transition genomic regions: the 34–37 Mb segment on chromosome 4 and the 10–13 Mb segment on chromosome 14 demonstrated an exception to the observed tendency. Four BAC clone-based probes, CH261-30O11, CH261-38A10 to the A compartment and CH261-124F10, CH261-109C8 to the B compartment, were selected for the 34–37 Mb region on chromosome 4 (GGA4) (Fig. 3dʹ). All four probes hybridized into the RNP-matrix of four adjacent lateral loops of different sizes regardless of whether they belonged to the A or B compartment (Fig. 3d–dʹʹ).
The GGA14 microchromosome contains two clusters of CNM repeats in the centromeric region corresponding to two large DAPI-positive chromomeres at the lampbrush chromosome stage [54] and an additional extended B compartment in the middle of the proximal part of the long arm (Fig. 5a–b). At the same time, three BAC clones from this B compartment mapped to three pairs of lateral loops, indicating active transcription of this region at the lampbrush chromosome stage [39]. In particular, the 748 kbp gene RBFOX1 (RNA Binding Fox-1 Homolog 1) present in this region is transcribed on a very long lateral loop [39] but is silent in chicken embryonic fibroblasts.
Here, we selected 9 additional and one previously mapped BAC clones for this region of GGA14–6 to the B compartment (CH261-177N7, CH261-36G10, CH261-179I1, CH261-119E18, CH261-78O7, CH261-99K17) and 4 to the A compartment (CH261-75C12, CH261-75E9, CH261-57E13, CH261-152F2) (10–13 Mb region, Fig. 5dʹ). Probes to the B compartment hybridized with the RNP matrix of two long adjacent lateral loops (Fig. 5d–dʹʹ). Probes to the A compartment also hybridized with the lateral loops, but despite the abundance of the genes in this region, the size of these loops was much smaller. This may be related both to the different lengths of the transcribed genes and to the intensity of their transcription at this stage of oogenesis. We assume that in the 10–13 Mb region of GGA14, the longest lateral loops are formed in the genomic regions containing long transcribed genes regardless of the compartment status.
Mapping genomic regions belonging to B compartmentsTo map the genomic regions belonging to the somatic B compartments, we chose a 185–186 Mb genomic segment that belongs to GGA1 region, dominated by B compartments interspersed with smaller A compartments (cluster #5) (Fig. 1a), and selected three neighboring BAC clones CH261-31K17, CH261-54J7, CH261-120O20 (Fig. 1fʹ). BAC clone-based probes CH261-31K17, CH261-54J7, and CH261-120O20 hybridized to three adjacent chromomeres and probe CH261-54J7 also hybridized with a pair of very short lateral loops, emanating from the central chromomere (Fig. 1f–fʹʹ). Thus, this cluster of DAPI-positive chromomeres on lampbrush chromosome 1 (cluster #5) (Fig. 1c) corresponds to the genomic region dominated by B compartments (Fig. 1a–b). Tiny lateral loops that emerge from these chromomeres are aligned with the small A compartment within this genomic region (Fig. 1f–fʹʹ). This example confirms the general correspondence between genomic regions forming B compartment in chicken embryonic fibroblasts and the large clusters of dense chromomeres with short lateral loops in lampbrush meiotic chromosomes.
In addition, the BAC probes that we earlier mapped on chicken lampbrush chromosome 1 to genomic loci from the two neighboring TADs belonging to the compartment B (146–148 Mb region) hybridize with one prominent chromomere, but not with lateral loops (Fig. 2 in [39]). Previously, microdissection and sequencing of a large dense chromomere from this region (149–154 Mb) showed that it is enriched with H3K9me3 and highly methylated gene-poor DNA corresponding to the somatic B compartment [35, 38]. In addition, among three BAC clones to the three subTADs from the compartment B (25–28 Mb region) on lampbrush chromosome 2 (Fig. 2a–c, region 2), two hybridized with chromomeres and one hybridized with the nascent RNA on a pair of lateral loops corresponding to the TSHD7A gene (Fig. 3 in [39]).
In conclusion, our FISH-mapping data additionally indicate that clusters of dense compact chromomeres carrying short lateral loops and enriched with repressive epigenetic modifications overlap with prominent B compartments in somatic cells, reflecting functional compartmentalization of the chicken genome. Next, we wondered if these compartments are constitutive or cell-type specific. Using the data available up to date, we demonstrate that these B compartments are constitutive between several chicken cell types: embryonic fibroblasts (CEF), mature erythrocytes (RBC), erythroblast cell line (HD3), DT40 cell line, granulosa cells of small white follicle (SWF), and liver cells (Additional file 1: Figure S2).
留言 (0)