
Remember me


Tinnitus is a prevalent disorder characterized by the perception of sound in the absence of an external sound source. Multiple causes of tinnitus have been described, such as hearing loss and temporomandibular joint disorders, but also stress and emotional factors seem to play an etiopathogenetic role (Langguth et al., 2013). Indeed, a clear association was found between both physical and psychological stress and the prevalence and severity of tinnitus (Hasson et al., 2011; Schaaf et al., 2014); however, the direction of causality in this association remains unclear (Pattyn et al., 2016; Pupi-Bakra and Pupi-Bakra, 2020). In both individuals with tinnitus and unaffected controls, noise causes subjective stress and activates the hypothalamic–pituitary–adrenal (HPA) axis, a major component of the neuroendocrine stress response (Hébert and Lupien, 2009). Vice versa, elevated levels of cortisol, the HPA axis’ end-product, can affect neuronal structures in the inner ear and as such pose a risk for the subsequent development of maladaptive central auditory responses and tinnitus (Mazurek et al., 2012; Knipper et al., 2013; Szczepek and Mazurek, 2021). Dysregulation of the HPA axis was indeed reported in tinnitus (Pattyn et al., 2016; Pupi-Bakra and Pupi-Bakra, 2020), although the number of studies on this topic is very limited. Baseline cortisol concentrations were shown to be higher in participants with tinnitus and high tinnitus-related distress in one study (Hébert et al., 2004). More recently, higher perceived tinnitus loudness was also reported to be associated with higher hair-cortisol levels (Basso et al., 2022). In contrast, in individuals with chronic tinnitus (≥6 months) a blunted cortisol response to psychosocial stress was found (Hébert and Lupien, 2007), as well as a hypersuppression of cortisol after administration of a low dose (0.5 mg) of dexamethasone, pointing to HPA axis hypofunction and to enhanced negative feedback to the HPA axis (Simoens and Hébert, 2012).
This attenuation of the HPA axis response has been described previously in response to different kinds of chronic stressors, with an inverse relationship between the time elapsed since the onset of a stressor and post-dexamethasone cortisol levels (Miller et al., 2007). It can accordingly be hypothesized that tinnitus acts as a chronic stressor, comparable to other chronic health problems (Malouff et al., 2011) with similar neuroendocrine profiles. Indeed, other functional syndromes such as chronic fatigue syndrome (CFS) are also associated with enhanced glucocorticoid feedback (Gaab et al., 2002; Van Den Eede et al., 2007; Papadopoulos and Cleare, 2011). Yet, the mechanism leading to this enhanced sensitivity to negative feedback (HPA axis hypersuppression) in response to stressors remains to be clarified.
Regarding this, recent research has provided greater insight into the potential role of epigenetic mechanisms: subtle molecular changes, sensitive to environmental stress, which modulate gene expression without altering the genetic code itself. For example, DNA methylation changes in genes encoding neurotrophic factors relevant to the auditory pathway have been demonstrated in chronic tinnitus (Orenay-Boyacioglu et al., 2019). Another well studied epigenetic phenomenon is the methylation of the glucocorticoid receptor gene (NR3C1), which plays a major role in HPA axis feedback. DNA methylation percentages of the 1F exon of the NR3C1 promotor region are reported to be lower in adults with CFS than they are in healthy counterparts (Vangeel et al., 2015), a finding that was later replicated in an independent CFS sample (Vangeel et al., 2018). Moreover, NR3C1-1F hypomethylation was associated with a stronger decline in plasma cortisol after administration of 0.5 mg dexamethasone, suggesting enhanced negative feedback to the HPA axis. In our chronic stress model of tinnitus, NR3C1-1F hypomethylation might also be responsible for the observed enhanced glucocorticoid feedback seen in this disorder. To our knowledge, this has never been investigated.
An important consideration is the frequent comorbidity of tinnitus and anxiety disorders (Pattyn et al., 2016; Oosterloo et al., 2021). Panic disorder, for example, which is characterized by recurrent unexpected panic attacks, is noticeably more prevalent in tinnitus populations (Mathias et al., 2011). Interestingly, there is an important clinical overlap between tinnitus and panic disorder: both conditions are accompanied by a heightened awareness of internal signals (Wallhäusser-Franke et al., 2013) and signs of autonomic arousal (Hiller et al., 1997); however, in a previous study by our group we found differential brain activity in tinnitus sufferers with comorbid panic disorder as compared to tinnitus-only and healthy controls (Pattyn et al., 2018). Furthermore, in contrast with the findings in tinnitus, a hyperreactivity of the HPA axis to novelty cues was reported in panic disorder (Abelson et al., 2007), as well as higher cortisol levels upon awakening (Vreeburg et al., 2010) and reduced negative feedback to the HPA axis (Schreiber et al., 1996); however, the related findings in panic disorder are somehow inconsistent because an intact negative feedback to the HPA axis has also been described (Ising et al., 2012) as well as a stronger suppression with lower cortisol levels after a high dose of dexamethasone than in healthy controls (Wichmann et al., 2018).
The HPA axis dysregulation in panic disorder is potentially driven by the typical anticipatory anxiety in-between panic attacks (Jakuszkowiak-Wojten et al., 2015), which can also be considered a chronic stressor. Several genes had distinct DNA methylation patterns in panic disorder compared to those observed for healthy controls (Kim et al., 2018; Ziegler et al., 2019; Petersen et al., 2020), including the gene encoding the corticotropin-releasing hormone receptor 1 (CRHR1) involved in the HPA axis response (Kim et al., 2018).
The primary objective of the current study was to examine associations between methylation of the NR3C1-1F and tinnitus in a case-control study comparing adults with chronic subjective tinnitus and healthy controls. Because an important fraction of individuals with tinnitus also report experiencing panic and panic attacks, our second objective was to test for a differential effect of panic on the DNA methylation patterns in our tinnitus sample.
Materials and methods Study populationPatients with subjective tinnitus and clinically significant comorbid anxiety were recruited at the Antwerp University Hospital from December 2015 to December 2017. Subjective tinnitus was diagnosed at the Department of Otorhinolaryngology when otorhinolaryngeal and neurological examination, as well as a complete audiological examination, failed to show a clear medical cause. To measure tinnitus-related distress, the Dutch version of the Tinnitus Functional Index (TFI) (Meikle et al., 2012; Dutch version validated by Tromp, 2014) was used, a self-report questionnaire consisting of 25 items that are scored on a 10-point Likert scale. Respondents with a score of 10 or more were considered to have significant psychological distress and were referred to our psychiatric outpatient unit within an integrated care framework. Here, patients were screened for potential recruitment in the study.
To delineate their anxiety symptoms, eligible participants completed the Beck Anxiety Inventory (BAI) (Beck et al., 1988; Dutch version validated by Vos, 2012), whose 21 items are to be scored on a four-point Likert scale. Panic attacks were assessed categorically according to the criteria of the Diagnostic and Statistical Manual of Mental Health Disorders Fifth Edition, American Psychiatric Association 2013, Washington, DC.
Because childhood trauma has consistently been reported to be correlated with NR3C1-1F methylation in mood disorders (Nöthling et al., 2020), it can be considered a potential confounder that should be taken into account when investigating NR3C1-1F methylation in panic disorder or panic attacks as well. To screen for a history of childhood trauma, the Netherlands Mental Health Survey and Incidence Study (NEMESIS) trauma interview (De Graaf et al., 2002) was administered. The NEMESIS trauma interview is a Dutch retrospective, semi-structured interview screening for the occurrence of childhood trauma (emotional neglect, psychological abuse, physical abuse and sexual abuse) before the age of 16. Participants are asked if and how often this has occurred. From the number of different types of experienced childhood trauma and their frequency (ranging from 0, “never,” to 2, “regularly to very often”), the childhood trauma index (CTI) is calculated (ranging from 0 to 8).
Age- and sex-matched healthy controls were recruited through advertising and mailings within the university and hospital in the same time period as the study group to minimize sampling bias due to seasonal effects.
To be eligible, participants had to be at least 18 years old. Exclusion criteria were a current diagnosis of major depressive or post-traumatic stress disorder, current substance abuse and a past or current diagnosis of bipolar or psychotic disorder as assessed with the Dutch version of the SCID-I/P interview (version 2.0) (First et al., 1997; Dutch version by van Groenestijn et al., 1999), because of the possible impact of these disorders on DNA methylation patterns. Pregnancy or lactation and treatment with corticosteroids or hormonal replacement therapy (with the exception of contraception) were also reasons for exclusion. Because our focus was on tinnitus as a chronic stressor, in our analyses we only included individuals who reported having experienced tinnitus for at least 6 months, the same cutoff Simoens and Hébert (2012) used in their study on enhanced suppression of the HPA axis after a dexamethasone challenge.
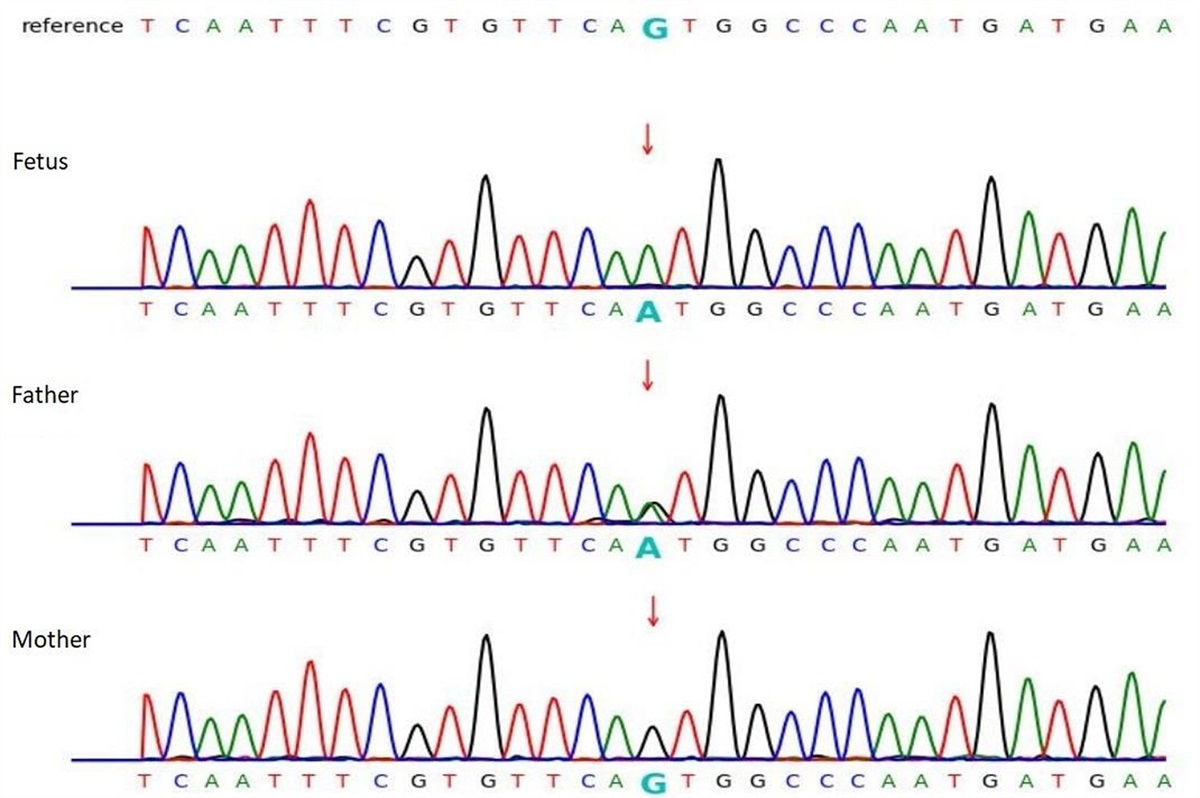
DNA methylation analysisPeripheral blood (2 mL) was collected in an EDTA tube (BD Vacutainer, Thermo Fisher Scientific, Massachusetts, USA). Samples were frozen immediately after collection and stored at −20 °C until processing. DNA was isolated from the peripheral blood samples at the Center for Medical Genetics (CMG) of Antwerp University Hospital using the QIAamp DNA Blood Mini Kit (Qiagen, Hilden, Germany). Bisulfite conversion of isolated DNA and subsequent quantitative methylation analyses were performed at the Genetic Research about Stress and Psychiatry research facility of the Division of Psychiatry of the Department of Neurosciences of the KU Leuven (Belgium). Due to the low DNA yield of the isolation process, the bisulfite conversion was carried out using the EZ-96 DNA Methylation-Direct Kit (Zymo Research, Irvine, California, USA), which has increased sensitivity. Next, a quantitative methylation analysis was run using pyrosequencing at eight CpG sites of the NR3C1 gene previously implicated in other populations exposed to chronic stressors such as a diagnosis of CFS (Vangeel et al., 2015,2018), a history of childhood trauma (Perroud et al., 2011) and in individuals coping with food and nutritional insecurity and depressive symptoms (Borçoi et al., 2021). First, PCR amplification of the NR3C1-1F promotor region (Fig. 1) was achieved using forward (5′-TTTGAAGTTTTTTTAGAGGG-3′) and reverse (5′-CCCCCAACTCCCCAAAAA-3′, with 5′ biotin essential for pyrosequencing analysis) primers (Perroud et al., 2011). Next, pyrosequencing was performed with the PyroMark Q24 (Qiagen) using the sequencing primer (5′- GAAAAGAAATTGGAG -3′). The percentages of DNA methylation for each CpG site were the outcomes used in the subsequent statistical analyses.
 Fig. 1: NR3C1 gene region surrounding the 1F exon. Underlined: exon 1F; red font in rectangle: PCR forward and reverse primers; blue font in oval: sequencing primer; green font in circles: analyzed CpG sites 1 through 8. Adapted from: Perroud et al. (2011).mRNA expression analysis
Fig. 1: NR3C1 gene region surrounding the 1F exon. Underlined: exon 1F; red font in rectangle: PCR forward and reverse primers; blue font in oval: sequencing primer; green font in circles: analyzed CpG sites 1 through 8. Adapted from: Perroud et al. (2011).mRNA expression analysis
Analysis of mRNA expression was also conducted at the CMG, Antwerp University Hospital. Total RNA was isolated from whole blood (5 mL) collected in PAXgene tubes (PreAnalytiX, Hombrechtikon, Switzerland) with the aid of the PAXgene Blood RNA kit (PreAnalytiX) and the DNA-free DNA Removal Kit (Thermo Fisher). Total RNA quality was checked by electrophoresis using the Experion RNA StdSens kit (Bio-Rad, California, USA). Given the relatively low RNA yield, isolated RNA was additionally concentrated using the RNA Clean & Concentrator kit (Zymo Research). This total RNA was then converted to cDNA using the SuperScript III First-strand Synthesis System (Thermo Fisher) with 300 ng of RNA as starting material. Forward (GATGACAACTTGACTTCTCTGG) and reverse (AGCTTACATCTGGTCTCATGCT) primers were developed and tested, yielding a mean amplification efficiency of approximately 100% [E = 2.040, SE(E) = 0.022] and adequate Cp values (<30) for cDNA concentrations of 100 ng/µl. Next, reference genes were selected using the geNorm Reference Gene Selection Kit and software (PrimerDesign, Camberley, UK). From this experiment TOP-1, UBC and ACTB were selected as the most stably expressed genes in our sample. Consequently, a two-step real-time PCR experiment was performed with the aid of the qPCR MasterMix Plus for SYBR Green I No ROX (Eurogentec, Seraing, Belgium).
Quality criteria for RNA analysisThe concentration and purity of the isolated total RNA samples was evaluated by electrophoresis using the Experion RNA StdSens kit (Bio-Rad). Samples were excluded if their RNA concentration was too low and if no 18S and 28S sample peaks were detected, either by the Experion software or upon visual inspection of the electropherogram. Next, samples were excluded if less than 120 ng of total RNA was available for subsequent cDNA conversion. As a final quality-control test, the presence of genomic DNA in the cDNA samples was checked by gel electrophoresis. Samples were excluded if they contained genomic DNA or if cDNA was undetectable upon visual inspection of the gel.
Statistical analysesIn the main analysis, differences in methylation across the eight CpG sites, which represents a within-subject repeated measurement, were modeled using linear mixed models. Methylation percentage was entered as the dependent variable, group and CpG site (categorical variable) as the fixed effect, with individual identifiers being added as random effects to account for the non-independence of observations within the same individual. The presence of position-dependent differences in methylation between the three groups was tested by adding an interaction term group × CpG site to the model and testing this interaction term for significance. In the absence of a significant group × CpG site interaction, a model with only the main effects of the CpG site and group was fitted. The significance of the main effect for the group indicates whether a difference in methylation was present across all CpG sites. If significant, a post-hoc analysis with Tukey correction for multiple testing was carried out to investigate with pairwise comparisons which groups were different from one another. The significance of the fixed effects was tested by means of an F-test with Kenward Roger correction. To fulfill the requirement for normality of the residuals and homoscedasticity, log-transformed values of the methylation percentage were entered as the dependent variable in the mixed model analysis.
Characteristics that were recorded only once in a participant, were tested for mean differences between the combined tinnitus group and the control group using the independent Student’s t-test. Differences between the tinnitus-only, tinnitus-with-panic attacks and control samples were tested using a one-way analysis of variance (ANOVA) followed by a post-hoc analysis with a Tukey correction for multiple testing. The association between the CpG values and the BAI was modeled using linear regression, and the correlation was expressed using Spearman’s correlation coefficient. Statistical analyses were performed using the software package R (The R Foundation for Statistical Computing, care of Institute for Statistics and Mathematics Wirtschaftsuniversität Wien; Welthandelsplatz 1; 1020 Vienna, Austria), version 3.5.1, and the packages lme4, pbkrtest and multcomp.
Ethical statementThe study was approved by the ethics committee of Antwerp University Hospital (file code 15/37/386) and conducted in accordance with the declaration of Helsinki. All eligible candidates gave their written informed consent prior to their participation in the study.
Results Sample characteristicsWe recruited a total of 22 patients with chronic tinnitus and 31 controls, whose characteristics can be found in Table 1. In the clinical group, half had tinnitus only, and half had tinnitus with comorbid panic attacks; see Table 2 and Supplementary Table 1, Supplemental digital content 1, https://links.lww.com/PG/A297 for the characteristics of these three groups.
Table 1 - Characteristics of the combined clinical group and the healthy control group (two-group comparison) Tinnitus (n = 22) Control (n = 31) Comparison Mean (SD) Mean (SD) F-statistic P value Age (in years) 43.55 (16.75) 41.84 (15.84) 0.142 0.71 BMI 26.06 (5.04) 24.17 (3.36) 2.303 0.14 TFI 46.34 (20.36) BAI 12.43 (12.48) 3.90 (5.56) 11.251 0.15E-2 HADS total 12.27 (6.93) 4.39 (3.76) 28.503 0.22E-5 HADS anxiety 6.86 (3.97) 2.84 (2.24) 22.113 0.20E-4 HADS depression 5.41 (4.10) 1.55 (2.26) 19.293 0.57E-4 Trauma score 1.91 (2.37) 0.90 (1.49) 3.5983 0.63E-1 Percentage Percentage Χ 2 P value Sex (women) 36.40 45.20 0.410 0.52 Smoking 4.50 12.90 1.052 0.31 Childhood trauma 45.45 32.26 0.475 0.49BAI, Beck Anxiety Inventory; HADS, Hospital Anxiety and Depression Scale; TFI, Tinnitus Functional Index.
BAI, Beck Anxiety Inventory; HADS, Hospital Anxiety and Depression Scale; TFI, Tinnitus Functional Index.
aOne-way ANOVA.
bTukey-corrected P value from post-hoc analysis. Effect sizes for pairwise comparisons are given in Supplementary Table 1, Supplemental digital content 1, https://links.lww.com/PG/A297.The tinnitus and control groups did not differ significantly regarding age, sex ratio, smoking and childhood trauma scores. The one-way ANOVA and post-hoc analysis showed that TFI scores were not significantly different for the two tinnitus groups (Tukey’s P = 0.73). The group difference in BAI scores was also significant, with the post-hoc analyses showing that this held for the tinnitus-with-panic attack group (P = 0.001) and not the tinnitus-only group (P = 0.27). There were no significant differences in BAI scores between the two tinnitus groups (P = 0.19) either, although, as expected, the mean BAI score was much higher in the tinnitus-with-panic attack group. The Hospital Anxiety and Depression Scale total score as well as the anxiety and depression scores for the controls differed significantly from those recorded for both tinnitus groups (P < 0.009 in all cases) but not between tinnitus groups (P = 0.55 in all cases).
DNA methylationOur two-group comparison (combined tinnitus vs. control samples) showed no overall main effect of affection state in the combined mixed model analysis including all CpG sites (P = 0.13). Moreover, none of the eight CpG sites showed a significant individual difference in NR3C1 methylation percentage (P > 0.05); however, our three-group comparison revealed that findings for the tinnitus-with-panic attack group were distinct from those obtained for the tinnitus-only and control groups. As can be seen in Fig. 2 and Supplementary Table 2, Supplemental digital content 2, https://links.lww.com/PG/A298 the difference in DNA methylation was relatively consistent and significant (P = 0.03) across CpG sites, with the post-hoc analysis indicating that methylation in the tinnitus-with-panic attack group was elevated. The difference with the control group was significant (P = 0.030 following Tukey correction for multiple testing), whereas the difference with the tinnitus-only group, though in the same order of magnitude, only showed a trend towards significance (P = 0.08). The differences in methylation patterns among the three groups were uniform across the CpG sites (P = 0.56, following the F-test for site × group interaction).
 Fig. 2:
Fig. 2: Methylation across the eight CpG sites in the three groups. The solid line connects the mean (log-transformed) methylation values from the eight CpG sides, for the three groups. Error bars indicate the 95% confidence interval around the mean.
All these associations remained unchanged upon accounting for childhood trauma using the CTI as a covariate in the regression model, albeit with more significant associations (P value for the main effect of group 0.012; Tukey-corrected P value for the pairwise difference between the tinnitus-with-panic attack and control groups 0.010); however, the effect of childhood trauma itself on CpG methylation was not significant (P = 0.33).
Because the difference in BMI among the subgroups approached significance (Supplementary Table 3, Supplemental digital content 3, https://links.lww.com/PG/A299P = 0.018), we added BMI as a covariate to the linear mixed model to account for this parameter in the association between methylation and tinnitus status. The addition of this covariate increased the significance of the association to a P value of 0.018.
Differences in mean methylation values were remarkably consistent across the eight CpG islands, with elevated methylation values in the tinnitus-with-panic attack group compared to the two other groups; however, testing each CpG site separately revealed that the difference was only significant for one of the CpG site (CpG7; P = 0.009), with two other islands showing a trend towards significance (CpG5 and 6; P = 0.080 and P = 0.095 respectively). Supplementary Table 2, Supplemental digital content 2, https://links.lww.com/PG/A298 shows the mean methylation levels for each of the eight CpG sites for each group.
Finally, given our finding that methylation percentages in the tinnitus-with-panic attack group were markedly more elevated than the rates observed for the tinnitus-only and healthy control groups, we performed additional analyses in the full sample to investigate whether high clinical anxiety (BAI total score) correlated with higher methylation percentages. We indeed found a significant positive correlation between methylation of the CpG7 island and the BAI total score (correlation = 0.408; P = 0.001). Supplementary Table 4, Supplemental digital content 4, https://links.lww.com/PG/A300 shows the correlation coefficients for the BAI outcomes and the CpG methylation for all eight CpG sites.
mRNA expressionEight samples were excluded from mRNA expression analyses because they did not meet the necessary quality criteria as described in the Methods section. Eventually, the mRNA expression analyses included samples of 10 participants in the tinnitus-only group, nine in the tinnitus-with-panic attack group and 27 of age- and sex-matched healthy controls. The characteristics of the mRNA expression analysis sample can be found in Supplementary Table 3, Supplemental digital content 3, https://links.lww.com/PG/A299.
There was no significant difference in mean NR3C1-1F expression between neither the combined tinnitus and control groups (F = 0.94, P = 0.34), nor between the two tinnitus groups and the healthy controls (F = 0.64, P = 0.63, one-way ANOVA), although the mean mRNA expression levels for the two tinnitus groups were increased with regards to the control group. As previously mentioned, DNA methylation is associated with the repression of transcription and thus reduced mRNA expression (Strachan and Read, 2011). To analyze whether, in tinnitus with concurrent panic, the deviant methylations are reflected in an altered expression pattern, we calculated the correlation between the NR3C1 expression and the methylation of the eight CpG sites around exon 1F in the three groups separately. In the healthy controls, the correlation between mRNA expression and NR3C1 promotor methylation percentages was very weak, while in both tinnitus groups, it was negative and more pronouncedly so in the tinnitus group with panic attacks (Fig. 3).
 Fig. 3:
Fig. 3: Correlograms of the (log-transformed) methylation percentages per CpG group with the Calibrated Normalized Relative Quantity (CNRQ) of NR3C1-1F mRNA expression (normalized with respect to the expression of the reference genes TOP-1, UBC and ACTB) and for (a) the healthy control group, (b) the tinnitus-only group and (c) the tinnitus group with panic attacks. The pairwise Spearman correlations are visualized by the color of the dots, with red and blue colors indicating a negative resp. positive Spearman correlation coefficient.
DiscussionIn this study, we studied differential DNA methylation in eight CpG sites around the 1F exon of the NR3C1, between adults with chronic subjective tinnitus with and without comorbid panic attacks and healthy control individuals. The combined group of tinnitus patients showed no significant differences from the healthy control group, but we found a consistently increased methylation across all eight CpG sites in the tinnitus group with comorbid panic attacks, with the most pronounced difference in CpG7. Interestingly, a significant positive correlation was found between methylation of the CpG7 site and the Beck Anxiety Inventory total score in the total sample; however, these differences in methylation were not paralleled by significant differences in expression.
The methylation patterns at the NR3C1 gene promotor region in our tinnitus sample did not differ significantly from the patterns found for the controls; however, there was a differential effect of panic attacks: the tinnitus-with-panic attack group did show consistently increased methylation, which was not observed in the tinnitus-only group, whose methylation patterns were similar to those of the controls. The lack of significant between-group differences in gene expression levels among the three
Comments (0)