3.1. Screening of Aflatoxins in ChocolatesAFB1 was the most frequently detected aflatoxin at the highest concentration, compared with AFB2, AFG1, and AFG2, in all chocolates. Accordingly, the highest mean concentration of AFB1 was detected in chocolate powder, at 1.12 µg/kg, exceeding the permissible limit according to Codex Standard (1 µg/kg) [
22]. On the other hand, our findings regarding AFs in all tested chocolate are within the allowable limits of the Brazilian Sanitary Surveillance Agency, which set limits of 10 μg/kg for cocoa beans and 5 μg/kg for cocoa products and chocolate sold in Brazil, for both ochratoxin A and total aflatoxins [
8].The contamination levels of AFG in powder and milk chocolates are significantly different (p
23], who found that AFG1 and G2 contents failed to be detected in all chocolate samples, except milk chocolate, where the concentration of AFG1 was 0.35 μg/kg.These results are relatively similar to those obtained by Copetti et al. [
24] who detected AFs in approximately 72% of milk chocolates and 100% of bitter and dark chocolate at concentration ranges of 0.11–1.65 and 0.04–0.91 μg/kg, respectively. In a study by Turcotte et al. [
25] the AFB1 was reported in 80% of dark chocolate and 70% of milk chocolate samples at values of 0.63 and 0.18 μg/kg, respectively. In Pakistan, Naz et al. [
26] reported that the contents of AFs in dark, milk, and bitter chocolates were 2.27, 1.31, and 0.97 µg/kg, respectively. In contrast, our findings are significantly higher than those reported by Kabak, [
23] who counted AFs in 19.6% of milk chocolate, 13.3% of bitter chocolate, and 8.70% of chocolate wafer samples at mean values of 0.839, 0.455, and 0.281 μg/kg, respectively. The authors confirmed that AFB1 was the highest detected AF in all examined chocolate samples.To date, there is no maximum threshold for AFs in cocoa and its products involving chocolate established by the European Regulation. The differences in AFs levels amongst examined chocolates may indicate the content and quality of cocoa solids. This finding may explain the high content of total Afs, 2.322 and 1.658 µg/kg, in cocoa powder and dark chocolate samples containing 100% and 35% cocoa solids, respectively. Although milk chocolate sample content was 29–35% cocoa solids [
22], the high content of total AFs (1.81 µg/kg) could be attributed to milk solids, based on the presence of AFs in milk products reported recently [
27]. The higher contamination levels of chocolates containing AFs may be attributable to the deficiency of quality control and quality assurance during the manufacturing, and unhygienic packaging. Currently, chocolate consumption among Egyptians, particularly children, is high. Consequently, quantifying the presence of mycotoxins in these products is a significant issue, and additional research is required to determine their mycotoxin concentrations. 3.2. Aflatoxin Adsorption Efficiency in Phosphate Buffer SolutionBy comparing the individual tested compounds, it was found that S. cerevisiae adsorbed the highest levels of AFs with adsorption vis residual levels of 71% vis 0.29 μg/mL and 73% vis 0.27 μg/mL at pH 3.0, and 71% vis 0.29 μg/mL at pH 6.8, after 2 and 4 h, respectively. This result is higher than those explained by Istiqomah et al. [
28], who found that the AFs binding activity of autoclaved S. cerevisiae B18 cells was 69.5%. Additionally, Rahaie et al. [
29] illustrated that the heat treatment of yeast cells increased their binding abilities to AFs by 56 ± 2%. A. charcoal adsorbed 52–55% (pH 3.0) and 52% (pH 6.8) of AFs versus 52–53% (pH 3.0) and 51% (pH 6.8) for L. rhamnosus, after 2 and 4 h, respectively. This finding indicates that L. rhamnosus-supplemented PBS demonstrated the highest levels of residual Afs, which were 0.48 and 0.47 μg/mL at pH 3.0, and 0.49 μg/mL at pH 6.8, for 2 and 4 h, respectively.
This investigation suggests that the adsorption efficiency depends on several reasons, e.g., physical, chemical, and biological properties of the adsorbent and the adsorbate (the food or beverage); concentration of the adsorbate in fluids; features of the liquid phase (e.g., pH, temperature) and the residence time. The tri-mix demonstrated the maximum AF adsorption, followed by the di-mix, due to the synergism among the tested adsorbents’ physical, chemical, ion-exchanging, and biological mechanisms. Accordingly, A. charcoal has a large surface area and pore volume, providing an excellent adsorption capability. It can sequester molecules, including mycotoxins, via both chemical and physical interactions; hence, it can be used as an adsorbent agent. A. charcoal was recognized to have binding efficiency of more than 90% with AFs.
Lactobacillus strains are Gram-positive bacteria, with a peptidoglycan layer in their cell wall and teichoic acid and polysaccharides (β-D-glucan) on the surface. The divalent cations can lead to alterations in the teichoic acid structure. Furthermore, the adsorption ability of lactobacilli is generally strain-dependent, due to the differences in the structure of biochemical components on the cell wall for most probiotic types [
30,
31].In this study, we found that all the PBS-tested compounds containing S. cerevisiae demonstrated the best AF adsorption. The adsorption capability mainly depended on yeast composition and mycotoxin. Earlier studies investigated that yeast cells had the ability to attach numerous molecules involving mycotoxins via the polysaccharides (glucans), mannoproteins, and lipids found on their cell wall surface [
32]. They can bind to the hydroxyl, ketone, and lactone groups of AFs by hydrogen bonds and van der Waals forces [
33]. Furthermore, the natural characteristics of mycotoxins play an integral role in the adsorption activities, such as polarization, solubility, size, shape, cation exchange capacity (CEC), acidity, and relative humidity [
34]. This finding agrees with Joannis-Cassan et al. [
35], who indicated that the yeast cell wall components played a crucial role in AFs binding by S. cerevisiae. Furthermore, S. cerevisiae was the most effective microorganism for binding AFs in PBS. Concerning the synergetic action achieved in this study, A. charcoal displays reversible physical adsorption for microorganisms in liquids without removal by simple desorption and permeability, permitting its usage as a bio preservative [
3]. A study by Ikegamai et al. [
36] revealed that the fermentation activities of S. cerevisiae reached 90% in the medium containing A. charcoal, compared to 70% in controls. Besides, attractive interactions between microorganisms and A. charcoal decreased the porosity and negative charges, enhancing the capacity of A. charcoal to adsorb AFs, which might be the chief cause of maximum adsorptive ability for the tri-mixed PBS solution.As hypothesized, the AF adsorption varied with the type and initial concentration of aflatoxin. Accordingly, AFB1 was the greatest adsorbed aflatoxin on A. charcoal + L. rhamnosus + S. cerevisiae-treated PBS, with residual values of 0.003 ± 0.01 μg/mL at pH 6.8 for 4 h, compared with the initial concentration of 0.24 ± 0.01 μg/mL. Furthermore, AFB2 achieved the best adsorption on A. charcoal + L. rhamnosus+ S. cerevisiae-treated buffer at pH 3 for 4 h compared to other Afs, with a residual level of 0 μg/mL of an initial concentration of 0.25 ± 0.01 μg/mL. Similar findings were obtained by El-Nezami et al. [
37] who found that L. rhamnosus strains successfully bound AFB1 and B2, rather than G1 and G2. In contrast, Liew et al. [
20] showed that the cell wall of lactobacilli showed the highest adsorption, at 97% for an AFB1 concentration of 6 µg/mL. Similarly, Hernandez-Mendoza et al. [
38] investigated the feasibility that teichoic acid contributes to AFB-binding by lactobacilli. The ability of bacterial cells to bind with AFs could be enhanced by chemical conditions, as bacterial cell treatment with acid facilitated the AFs’ physical binding to the bacterium’s molecular components, especially at the cell wall level.
With respect to AFG2, it was the most efficiently absorbed aflatoxin by S. cerevisiae, and A. charcoal + S. cerevisiae treated PBS compared to other AFs with residual values of 0.07 ± 0.01 and 0.02 μg/mL, respectively. Moreover, AFG1 and G2 showed a relatively lower residual level (0.005 ± 0.002 μg/mL) on A. charcoal + L. rhamnosus + S. cerevisiae tri-mixed PBS when compared with AFB1 and B2 on the same buffer. All AFs showed the same residual values on A. charcoal + L. rhamnosus-treated PBS to be 0.04 ± 0.01 μg/mL.
The adsorption efficiency in this study is higher than those recorded by Shetty et al. [
39], who examined the adsorption of AFB1 by 18 species of Saccharomyces at AFB1 concentrations of 1, 5, and 20 µg/mL. The authors reported that the yeast cells adsorbed 69.1% of the AFB1 at 1 µg/mL, 41.0% at 5 µg/mL of AFB1, and 34.0% at 20 µg/mL of AFB1. In this study, an increase in adsorption was achieved at a reduced initial concentration of each AF, around 0.25 ± 0.01 μg/mL, so the adsorption levels were relatively high. These findings are compatible with Joannis-Cassan et al. [
35] who reported that the range of adsorption was 2.5% to 49.3%, based on the AFB1 concentration and the adsorbent type. Similarly, Gallo and Masoero [
40] reported that the adsorption varied from 32% to 54%, with an initial AFB1 concentration of 0.82 µg/mL.By modifying surface charges and adsorbent–adsorbate reactions, pH substantially affects a liquid medium’s adsorption efficiency. Hence, the adsorption ability of all adsorbents for the AFs was evaluated at 2 pH levels of 3.0 and 6.8. In this regard, we found that the only treatment of PBS either by A. charcoal or L. rhamnosus, or S. cerevisiae achieved the greatest adsorption, ranging from 52% to 73% at acidic pH (3.0). In a study by Joannis-Cassan et al. [
35] they demonstrated that acidic pH (3.0) permitted adequate adsorption of the mycotoxins in the buffer. Exchangeable cations are involved in the binding mechanisms of AFs, which can be enhanced after acidulating the adsorbent surface.In contrast, the di-mixed treatment of PBS by A. charcoal + L. rhamnosus, A. charcoal + S. cerevisiae, or L. rhamnosus + S. cerevisiae demonstrated the best adsorption, varying from 80% to 91% at pH 6.8. The tri-mixed PBS by A. charcoal + L. rhamnosus + S. cerevisiae had the overall maximum adsorption of 97.7% and 99.70% at pH 6.8. These results indicate that the adsorption of AFB1 and AFB2 by the combined adsorbents was pH-independent, whereas adsorption of AFG1 and AFG2 increased to a certain extent at neutral pH (6.8), as evidenced by the net surface charges on adsorbents and the charge on AFs molecules. Similar findings were reported by Rasheed et al. [
41] who found no evident variation in the adsorption for AFB1 and AFB2 in buffer at pH 7, while for AFG1 and AFG2, adsorption increased enormously. Moreover, this result agrees with Tejada et al. [
42] who noticed that the highest adsorption of A. charcoal reached a value of 75.4% in a solution at pH 6.In the present research, we used two time intervals, and the adsorption efficiency of AFs by all treated buffers was more significant at 4 h than at 2 h. This result is compatible with Rahaie et al. [
29], who confirmed that the adsorption capability of AFs in a buffer may occur at 2 h to 3 h of treatment. The variation in the incubation time could be the factor that influences the adsorption efficiency of AFs. In the current study, the incubation times of ranging from 2–4 h were used to reflect the transit time in human GIT. In addition, the absorption of AFs mainly occurs in the small intestine [
19]. Generally, incubation time plays a part in the adsorption activity.

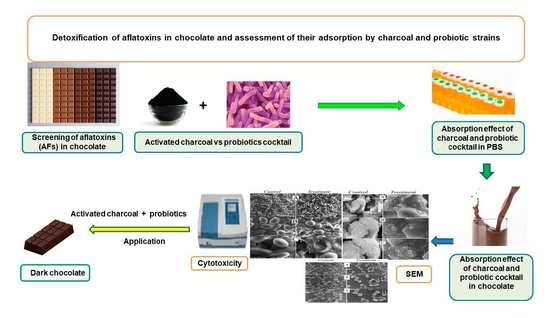


Comments (0)