1. IntroductionIn the last few decades, improvements in the survival rate of preterm infants have been observed [
1]. The management and prevention of complications arising from preterm birth and immature organs, such as necrotizing enterocolitis (NEC), feeding intolerance (FI) and sepsis, are crucial to the developmental outcome of these infants. Prematurity is a consistent independent risk factor of NEC, and NEC incidence is known to increase with decreasing birth weight and gestational age (GA) [
2,
3]. However, prematurity itself cannot entirely explain the pathogenesis of NEC/FI because some infants born at higher GA may also manifest NEC/FI. Due to the multifactorial and complex etiology, the pathogenesis of NEC has remained elusive and controversial until now [
4,
5,
6]. The intestinal microbiota performs several functions, including maturation of the immune system, regulating immune response, protection against the invasion of opportunistic pathogens, and mediating hormonal regulation [
7]. Prior to birth, infants are isolated from exposure to microorganisms in the environment [
8], and fetal meconium has been reported to be without any detectable microbiota before birth [
9]. Other studies suggest that the environment in utero is not sterile, and therefore the first passed meconium may somewhat mirror the in utero microbial environment [
10,
11]. Nevertheless, perinatal exposures to microbes lead to transient or long-term colonization. Therefore, perinatal events such as the method of delivery affect the types of microbes that infants are exposed to, resulting in an establishment of different types of gut microbiomes [
12,
13,
14].The preterm gut microbiome undergoes early disruption before achieving bacterial maturation. Gestational age at birth, representing the timing of first microbial exposure, is known to be significantly associated with the microbial composition including biomass and diversity of meconium in preterm infants [
14,
15]. In fact, according to a study examining the relationship of NEC to GA, events occurring at or soon after birth was found to be important in NEC development [
16]. The development of NEC highly involves gut mucosal immunity, comprising intestinal cell signaling and gut microbiota [
17]. The immature intestine of premature infants may be in a hyperactive state with increased inflammatory responses and impairment of intestinal perfusion [
18]. Thus, the timing and state of the preterm infants’ gut can influence the composition of the gut microbiome leading to the development of NEC.
In order to understand how the composition of the gut microbiome in the immediate perinatal period may affect the clinical course of preterm and term infants, the primary aim of this study was to investigate the composition of the first passed meconium microbiome in Korean preterm and term infants. The secondary aim was to compare the meconium microbiomes of preterm and term infants that later developed NEC/FI versus those that did not develop NEC/FI.
2. Materials and Methods 2.1. Study Population and Sample Collection
This was a prospective study carried out at a neonatal intensive care unit (NICU) of a tertiary referral university hospital located in Seoul, Korea. Preterm and term infants admitted to the NICU during May 2021 to February 2022 were included as study participants. The inclusion criteria were as follows: (1) first meconium stool sample obtained within 72 hours of birth during the study period, (2) admitted to the NICU regardless of the GA at birth, (3) transferred to the NICU immediately after birth for any symptoms requiring intensive care, and (4) obtained informed consent from the legal guardians. Exclusion criteria were as follows: (1) transferred to the NICU after being admitted to the nursery or another hospital, and (2) first meconium stool passed 72 hours or more after birth.
Immediately upon passage, 1–2 mL (maximum 3 mL) of the infant’s first passed meconium was collected and stored at −20 °C. The specimens were then transferred to a deep freezer set at −80 °C until DNA was extracted for microbiome analyses. The clinical course and progression of the infants included in the study were surveyed until death or discharge. Demographic and clinical data, including GA, birth weight, history of sepsis, and postnatal hospital outcomes were obtained. The patients were divided into two groups: NEC/FI group comprised 26 (24.1%) infants that were diagnosed with NEC/FI during the course of prospective monitoring, and the control group consisting of 82 infants (75.9%) that did not present with any GI (gastrointestinal) symptoms during the NICU admission period.
The study was approved by the Ethics Committee of Seoul St. Mary’s Hospital, The Catholic University of Seoul, Korea (IRB no. KC21TISI0329). All legal guardians of the study participants gave approval and signed informed consent forms for study participation.
2.2. Fecal Microbiome Analysis; DNA Extraction and AmplificationDNA was extracted using TIANLONG ®-nucleic acid extraction kit (for stool DNA/RNA extraction) (TianLong Science and Technology Co., Xi’an, China) then determined concentration and purity using nanodrop spectrophotometer. The target region (V3-V4) was amplified using PCRBIO VeriFi Mix (PCR Biosystems®, London, UK) and 16S-amplicon primer (Macrogen®, Seoul, Republic of Korea) at 95 °C for 3 min hot start followed by 25 cycles of 95 °C for 30 s and 55 °C for 30 s, 72 °C for 30 s with a final elongation step of 72 °C for 5 min. The V3-V4 target region reverse primer (5'TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGCCTACGGGNGGCW-GCAG3’) and forward primer (5'GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGGACT-ACHVGGGTATCTAATCC3’) were used according to the guideline ‘Preparing 16S Ribosomal RNA Gene Amplicons for the Illumina MiSeq System’ [
19].
The amplified DNA subsequently were purified automatically in Nucleic Acid Extractor (TianLong Science and Technology Co.) using MagListo™ PCR/Gel Purification Kit (Bioneer®, Daejon, Republic of Korea), then the size and concentration were determined using Qsep100 (Bioptic®, Changzhou City, China). Purified samples were then standardized to 8 ng per reaction (total reaction volume of 20 µL) and Index PCR was performed to attach in dual indices and Illumina sequencing using PCRBIO VeriFi Mix (PCR Biosystems®, London, UK) and Nextera ® Index kit V2 Set A (Illumina®, San Diego, CA, USA) at 95 °C for 3 min hotstart followed by 8 cycles of 95 °C for 30s and 55 °C for 30 s, 72 °C for 30 s with a final elongation step of 72 °C for 5 min.
After indexing, the final library was cleaned up using MagListo™ PCR/Gel Purification Kit (Bioneer®) before quantification. DNA size and concentration were again measured using Qsep100 (Bioptic®) and Qbit flex fluorometer (Invitrogen®). After Qbit and Qsep measurements were completed, the mixing volume value was calculated then the PCR product was pooled into a microtube. After mixing, the final pooled library concentration and size were checked using Qbit and Qsep. The counts per reaction for each microbiome species was recorded.
2.3. Sequencing and Data AnalysisLibraries were prepared using MiSeq ® Reagent Kit V3 600 cycles kit (Illumina®), pooled libraries were denatured with NaOH 0.2 N, then diluted with hybridization buffer before sequencing, PhiX was used as an internal control for each run. DNA were pooled and sequenced on an Illumina Miseq platform according to the manufacturer’s standard instruction, sequencing data of bacterial variability sites (V3-V4) were analyzed with 16S metagenomics app. Analysis was performed on Quantitative Insights into Microbial Ecology 2 (QIIME2; version 2020.2 and 2020.6) [
20]. DNA reads under 200 bp were omitted from the taxonomic analysis. Reads were then demultiplexed and denoised with DADA2 [
21]. Denoised reads were trimmed at 15 and truncated at 260, and chimeric reads were filtered out, resulting in a total of 3,994,640 processed reads ready for further analyses. R package decontam (version 1.8.0) was used to filter out environmental contaminants from each sample type [
22]. 2.4. DefinitionsNEC was defined as stage II or above, according to the modified Bell’s staging classification grade [
23], which includes one or more of the following clinical signs: bilious, gastric aspirator emesis, abdominal distention, or occult or gross blood in stool. This classification also includes one or more of the following radiographic findings: pneumatosis intestinalis, hepatobiliary gas, or pneumoperitoneum. Therapeutic decisions were based on clinical staging. FI was defined as persistent gastric aspirates of >50% of the feed volume with or without increased abdominal girth in the absence of culture-positive sepsis or radiographic evidence of NEC for 48 hours [
24], more than 3 times a day which did not allow the advancement of feeding > 10–20 mL/kg/day. Respiratory distress syndrome (RDS) was diagnosed based on both clinical and radiographic findings. Bronchopulmonary dysplasia was defined as use of oxygen ≥ 0.21 at 36 weeks.Preterm infants were defined as infants born
25]. Small for gestational age was defined as a neonate born with birthweights below the 2.5. Statistical Analysis
For descriptive statistics with demographic data and concentration and continuous variables were presented as means and standard deviation (SD) while categorical variables were presented as percentages and frequencies. For inferential statistics, continuous variables were compared using the t test, one-way ANOVA or Wilcoxon rank sum test depending on the normality of the variable being tested. More specifically, we compared the microbiome concentrations in the NEC group versus controls using the student’s t test (or Welch–Satterthwaite t test when the variance in the two groups was unequal). The logistic regression analysis was used to find the P for trend. The alpha diversity was calculated using Shannon’s diversity index, and beta diversity was plotted using Principal Coordinate Analysis of Bray–Curtis dissimilarity. Differences in categorical variables are compared using the chi-square or Fisher’s exact test. A univariate and multivariate analyses was carried out on all possible perinatal factors associated with NEC/FI. A p value of <0.05 was considered statistically significant.
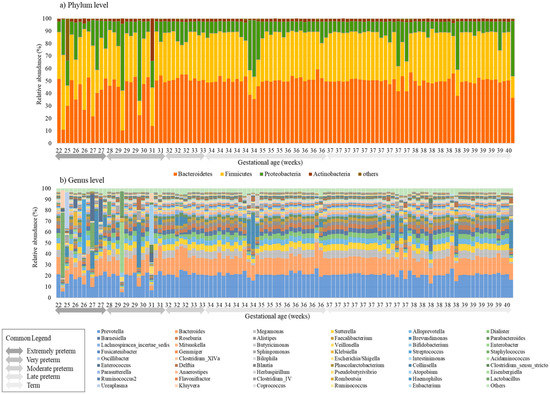
4. DiscussionThe gut microbiota is a group of microorganisms, mainly bacteria, yeast, fungi, bacteriophages, and other viruses, as well as protozoa and archaea, which form a complex ecosystem in the human GI tract [
27,
28,
29]. Meconium is often defined as the first stool passed within 48 h from birth. Neonatal microbiota starts diversifying quickly after birth, and compared to adults or older children, infant microbiota is known to have lower diversity as well as an unstable and highly dynamic microbiota structure [
30]. This present study investigated the gut microbiome of 108 preterm and term infants' first passed meconium. The microbiomes of the meconium of preterm infants up to 30 weeks of GA showed lower alpha diversities and disruption in the percentages of the core microbiome in both the phylum and genus level, compared to infants born at a higher GA portraying a difference in the perinatal gut microbiome of preterm and term infants. The two phyla, Bacteroidetes and Firmicutes, constituted majority of the microbiota of the neonatal meconium, and as the GA increased, a significant increase in the relative abundance of Bacteroidetes and decrease in Proteobacteria was observed. A study from Germany also showed that GA was significantly associated with the composition of meconium from very preterm infants, and the most abundant phyla included Firmicutes, Bacteriodetes, Proteobacteria, and Actinobacteria [
14]. Our study showed that in the genus level, Prevotella and Bacteroides were the predominant species, with Prevotella’s relative abundance accounting for 20-30% of the gut microbiome. Whereas in healthy adults, the two main phyla—Firmicutes and Bacteroidetes—constitute over 90% of the gut microbiota, followed by Actinobacteria and Proteobacteria [
31,
32]. Upon delivery, infants are exposed for the first time to a variety of microbes from various sources which lead to an establishment of the infants’ gut microbiome. Recent studies have shown that the predominant early colonizers of infant’s gut are maternal fecal bacteria, mainly Bifidobacterium and Bacteroides, and Clostridium [
27,
33,
34,
35]. Studies have also demonstrated that Bacteroides are associated with increased gut diversity and faster intestine maturation and natural childbirth has been shown to be significantly related to microorganisms reflecting the mother’s vaginal flora, such as Bacteroides, Lactobacillus, and Prevotella [
36,
37]. In the first passed microbiome of neonates in our study, Prevotella and Bacteroides were the most prevalent, accounting for about 50% of the core microbiome. However, because 83.3% (n = 90) were born via caesarean section, Lactobacillus was not a predominant genus in the meconium of the infants in our study. One of the most devastating gastrointestinal emergencies and a major cause of mortality in very low birth weight infants (VLBWI) is NEC. Furthermore, FI is also a frequently faced problem in many neonatal intensive care units (NICUs). Prenatal and postnatal factors, such as placental insufficiency, chorioamnionitis, gut ischemia, altered bacterial colonization, viruses, and blood transfusions, presumably disrupt the mucosal barrier, which may trigger inflammatory reactions in the immature intestines of preterm infants [
4]. This study investigated whether a difference can be observed in the microbiome of meconium of infants that are eventually diagnosed with NEC. This study found that compared to infants that are discharged from the NICU without any gastrointestinal symptoms, infants that were diagnosed with NEC/FI had significantly lower alpha diversities in their first meconium. Furthermore, a significantly lower abundance of Bacteroidetes, and higher abundance of Firmicutes and other microbes were observed. Firmicutes, Streptococcaceae, and Enterococcaceae are known to predominate in adults with lower diversity gut microbiota [
38]. Lower diversity gut microbiota is also known to be drivers of many diseases such as inflammatory bowel disease, acute diarrheal disease, C. difficile infection, and observed in cancer patients [
38,
39,
40,
41]. This shows that infants with NEC/FI have lower diversity dysbiosis, and this may have an important role for triggering inflammation leading to NEC/FI. In the study on meconium microbiome and its relation to neonatal growth and head circumference catch-up in preterm infants, Polynucleobacter, Gp1, and Prevotella appeared in greater abundance in meconium of preterm infants with adequate birth weight for GA [
42]. In our study, we found that the composition of Prevotella was positively correlated with GA, suggesting that the abundance of Prevotella has an important role in the growth and development of preterm infants. The recent study conducted by Liu et al. on early gut microbiota in very low and extremely low birth weight preterm infants with feeding intolerance showed that the meconium samples of the FI group had higher proportions of γ-proteobacteria and Escherichia-Shigella and a lower proportion of Bacteroides compared with the those that did not have FI [
43]. Although this was not corroborated in our study, our study found a lower proportion of Bacteroides in preterm infants with lower GA, suggesting that Bacteroides is a part of the healthy gut microbiome. Perturbation of the gut microbiota may promote overgrowth of pathobionts increasing the risk of infections and inflammation in the gut [
44]. In a study on infants born via caesarean section, a high-level colonization by opportunistic pathogens was observed, including Enterococcus, Enterobacter, and Klebsiella [
12]. In this study, the average counts in the NEC/FI group for the following pathobionts were significantly higher: Klebsiella, Streptococcus, Staphylococcus, Enterococcus, and Ureaplasma, and Kluyvera. The first moments of life are known to contribute to the formation of an NEC-associated microbiota [
45], however, our study shows that the initiation in the formation of an NEC associated microbiota may even occur prior to events following birth.In this study, there were six cases of placental abruption and 14 cases of PROM, meaning that these infants may have been exposed to the vaginal microbiota in utero, and subsequently affecting the results of the gut microbiome in these subjects. Due to the small number of subjects, further analyses were difficult. However, a study on 8 preterm neonates in Indonesia reported that PROM and mother’s diet influenced the meconium microbiome [
46]. Further studies including a larger cohort of preterm infants are needed to make any conclusions on the impact of early exposure to vaginal microbiomes in utero and colonization and establishment of the infants’ gut microbiome. When compared with bacteria, the healthy human gut mycobiome is lower in biomass compared to bacteriome biomass. However, the roles of mycobiomes are increasingly recognized as important, whether it be beneficial or harmful. The gut mycobiome is known to be dominated by Saccharomyces, Malassezia, and Candida [
47], and these pathobionts may potentially be harmful to preterm infants [
48]. Further studies are needed to characterize the composition of mycobiomes in preterm infants to better understand its role in the immature gut. This study had a few limitations the sample size of the study groups was relatively small. However, to date, this study contains one of the largest cohort of neonates, from GA 22 to 40. Secondly, although not conclusive, studies have shown evidence of sex related difference in the gut microbiome [
49]. Other studies do not support this [
50]. In our study, 88.9% (n = 96) patient were male, therefore there may have been selection bias genders-wise. There is a lack of studies on the influence that gender has in the gut microbial composition of infants, and further studies are warranted. Finally, this study lacked a longitudinal analysis of changes in the gut microbiome of neonates after birth. However, the focus of our study was investigating the microbiome of the first meconium passed by the infant immediately after birth reflecting the perinatal gut microbiome of the infant.
In summary, this study showed that low diversity dysbiosis was observed in the initial gut microbiomes of infants born premature, below 30 weeks of GA, and an increasing abundance of Bacteroidetes was observed as gestation age increased. Infants with NEC/FI had significantly lower alpha diversities, and a significantly higher counts of pathobionts such as Klebsiella, Streptococcus, Staphylococcus, Enterococcus, Ureaplasma, and Kluyvera were observed. To conclude, differences were observed in the composition of the first passed meconium in preterm and term infants that later developed NEC/FI compared to those that did not. These findings fuel the need to understand disease pathogenesis and develop novel and significant microbiota for earlier therapeutic and preventative strategies for vulnerable preterm babies in further research.

Comments (0)