














Remember me

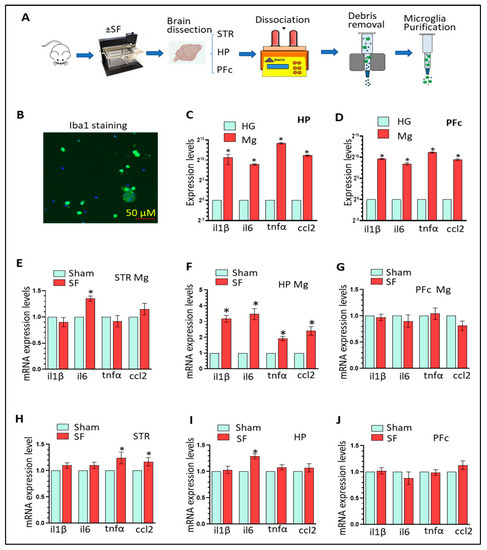

Figure 1. The cellularity of the local draining lymph node changed following dendritic cell (DC)-based vaccination.Ex vivo-generated DC-based vaccines were administered into the hind footpads of female C57Bl/6 mice (n = 2–7 per treatment), and then the popliteal lymph nodes were examined six-, 12-, 24-, 48-, and 120 h post-immunization to evaluate changes in cellularity. The number and percentage of natural killer (NK) and T cells in the lymph nodes were quantified at each time point following DC immunization using flow cytometry and compared with lymph nodes from control mice that received injections of phosphate-buffered saline (PBS). Statistical analysis was performed using a two-way analysis of variance; p-value < 0.05 (*), p-value < 0.001 (***), p-value < 0.0001 (****). Graphs show the means and the standard deviation with each dot representing an individual mouse.
Figure 1. The cellularity of the local draining lymph node changed following dendritic cell (DC)-based vaccination.Ex vivo-generated DC-based vaccines were administered into the hind footpads of female C57Bl/6 mice (n = 2–7 per treatment), and then the popliteal lymph nodes were examined six-, 12-, 24-, 48-, and 120 h post-immunization to evaluate changes in cellularity. The number and percentage of natural killer (NK) and T cells in the lymph nodes were quantified at each time point following DC immunization using flow cytometry and compared with lymph nodes from control mice that received injections of phosphate-buffered saline (PBS). Statistical analysis was performed using a two-way analysis of variance; p-value < 0.05 (*), p-value < 0.001 (***), p-value < 0.0001 (****). Graphs show the means and the standard deviation with each dot representing an individual mouse.
Figure 2. Rapid recruitment of a Ly6G+ population to the local draining lymph node following dendritic cell (DC) vaccination.Ex vivo-generated DC vaccines were injected into the hind footpads of female C57BL/6 mice (n = 2–6 per treatment), and the local draining lymph node was examined for Ly6G+ cells. Ly6G+ cells were assessed by flow cytometry six, 12, 24, 48, and 120 h post-DC immunization. The number and percentage of Ly6G+ cells in the lymph node of DC-immunized mice were compared with phosphate-buffered saline (PBS)-injected control mice. Significance was determined statistically using a two-way analysis of variance.; p-value < 0.05 (*), p-value < 0.0001 (****). Graphs show the mean and the standard deviation with each dot representing an individual mouse.
Figure 2. Rapid recruitment of a Ly6G+ population to the local draining lymph node following dendritic cell (DC) vaccination.Ex vivo-generated DC vaccines were injected into the hind footpads of female C57BL/6 mice (n = 2–6 per treatment), and the local draining lymph node was examined for Ly6G+ cells. Ly6G+ cells were assessed by flow cytometry six, 12, 24, 48, and 120 h post-DC immunization. The number and percentage of Ly6G+ cells in the lymph node of DC-immunized mice were compared with phosphate-buffered saline (PBS)-injected control mice. Significance was determined statistically using a two-way analysis of variance.; p-value < 0.05 (*), p-value < 0.0001 (****). Graphs show the mean and the standard deviation with each dot representing an individual mouse.
Figure 3. Ly6G+ cell recruitment was limited to the vaccine-draining lymph node. Dendritic cell (DC) vaccines were injected into one hind footpad of each female C57BL/6 mouse and phosphate-buffered saline (PBS) into the other hind footpad. Both popliteal lymph nodes were examined for Ly6G+ populations 12 h post-immunization, one representing the PBS-draining lymph node (left panel) and the other as a local vaccine-draining lymph node (middle panel). The inguinal lymph node on the vaccine-administered side of the mouse was also examined as a non-vaccine-draining lymph node (right panel). Representative flow cytometry dot plots of live cells in the lymph nodes are shown.
Figure 3. Ly6G+ cell recruitment was limited to the vaccine-draining lymph node. Dendritic cell (DC) vaccines were injected into one hind footpad of each female C57BL/6 mouse and phosphate-buffered saline (PBS) into the other hind footpad. Both popliteal lymph nodes were examined for Ly6G+ populations 12 h post-immunization, one representing the PBS-draining lymph node (left panel) and the other as a local vaccine-draining lymph node (middle panel). The inguinal lymph node on the vaccine-administered side of the mouse was also examined as a non-vaccine-draining lymph node (right panel). Representative flow cytometry dot plots of live cells in the lymph nodes are shown.
Figure 4. Neutrophils were recruited to the dendritic cell (DC) vaccine-draining lymph node. DC vaccines were generated ex vivo and administered into the hind footpads of female C57BL/6 mice (n = 2–4 per treatment). The vaccine-draining popliteal lymph nodes (labeled “DC”) were examined for neutrophils (Ly6G+ CD11b+) six-, 24-, and 48 h following DC vaccination and were compared with non-draining inguinal lymph nodes from the vaccinated side of mice (“DC non-draining”), and phosphate-buffered saline- draining lymph nodes (“PBS”). Statistical analysis was performed using a two-way analysis of variance; p-value < 0.05 (*), p-value < 0.001 (***), p-value < 0.0001 (****). Graphs show the means and the standard deviation with each dot representing an individual mouse.
Figure 4. Neutrophils were recruited to the dendritic cell (DC) vaccine-draining lymph node. DC vaccines were generated ex vivo and administered into the hind footpads of female C57BL/6 mice (n = 2–4 per treatment). The vaccine-draining popliteal lymph nodes (labeled “DC”) were examined for neutrophils (Ly6G+ CD11b+) six-, 24-, and 48 h following DC vaccination and were compared with non-draining inguinal lymph nodes from the vaccinated side of mice (“DC non-draining”), and phosphate-buffered saline- draining lymph nodes (“PBS”). Statistical analysis was performed using a two-way analysis of variance; p-value < 0.05 (*), p-value < 0.001 (***), p-value < 0.0001 (****). Graphs show the means and the standard deviation with each dot representing an individual mouse.
Figure 5. Neutrophil accumulation in the draining lymph nodes required maturation of dendritic cells (DCs). DC vaccines were prepared using DCs cultured ex vivo. The DCs were matured using lipopolysaccharide (LPS) as a source of pathogen-associated molecular pattern signaling that induces activation and maturation of DCs, and then they were loaded with antigens. DC cultures were prepared for immunization using different iterations of the manufacturing protocol. These included DCs stimulated with LPS only (DC LPS), DCs pulsed with antigens only (DC Ag), DCs pulsed with antigens and stimulated with LPS (DC Ag LPS), and DCs without LPS treatment or antigen-pulsing, which were termed “DC empty”. The number and percentage of neutrophils (Ly6G+ CD11b+) in the draining lymph nodes of the female C57BL/6 mice were examined one day following inoculations using flow cytometry. The data were analyzed for significance using a one-way analysis of variance; p-value < 0.005 (**), p-value < 0.001 (***), p-value < 0.0001 (****). Graphs show the means and the standard deviation with each dot representing an individual mouse.
Figure 5. Neutrophil accumulation in the draining lymph nodes required maturation of dendritic cells (DCs). DC vaccines were prepared using DCs cultured ex vivo. The DCs were matured using lipopolysaccharide (LPS) as a source of pathogen-associated molecular pattern signaling that induces activation and maturation of DCs, and then they were loaded with antigens. DC cultures were prepared for immunization using different iterations of the manufacturing protocol. These included DCs stimulated with LPS only (DC LPS), DCs pulsed with antigens only (DC Ag), DCs pulsed with antigens and stimulated with LPS (DC Ag LPS), and DCs without LPS treatment or antigen-pulsing, which were termed “DC empty”. The number and percentage of neutrophils (Ly6G+ CD11b+) in the draining lymph nodes of the female C57BL/6 mice were examined one day following inoculations using flow cytometry. The data were analyzed for significance using a one-way analysis of variance; p-value < 0.005 (**), p-value < 0.001 (***), p-value < 0.0001 (****). Graphs show the means and the standard deviation with each dot representing an individual mouse.
Figure 6. Soluble factor(s) produced by mature dendritic cells (DCs) were capable of recruiting neutrophils to the local draining lymph node. DCs were cultured ex vivo and half of the culture was matured using lipopolysaccharide, and the other half remained in an immature state. The DCs were plated in media and left overnight in an incubator at 37 °C. The supernatants of the immature and mature DCs were collected and injected into the hind footpads of female C57BL/6 mice (n = 4–7 per treatment), and media only was injected into negative control mice. Four hours post-injection, the draining lymph nodes were examined for neutrophils (Ly6G+ CD11b+). Significant differences were examined using a one-way analysis of variance; p-value > 0.0001 (****). Graphs show the means and the standard deviation with each dot representing an individual mouse.
Figure 6. Soluble factor(s) produced by mature dendritic cells (DCs) were capable of recruiting neutrophils to the local draining lymph node. DCs were cultured ex vivo and half of the culture was matured using lipopolysaccharide, and the other half remained in an immature state. The DCs were plated in media and left overnight in an incubator at 37 °C. The supernatants of the immature and mature DCs were collected and injected into the hind footpads of female C57BL/6 mice (n = 4–7 per treatment), and media only was injected into negative control mice. Four hours post-injection, the draining lymph nodes were examined for neutrophils (Ly6G+ CD11b+). Significant differences were examined using a one-way analysis of variance; p-value > 0.0001 (****). Graphs show the means and the standard deviation with each dot representing an individual mouse.
Figure 7. Neutrophils influenced dendritic cell (DC) vaccine recovery in the footpad and draining lymph node one day post-immunization. DC vaccines were prepared ex vivo and labelled with the fluorescent dye, carboxyfluorescein succinimidyl ester (CFSE). Female C57BL/6 mice (n = 8–16 per treatment) were treated with an anti-Ly6G antibody one day prior to and on the day of DC immunization to deplete neutrophils from the mice. The footpads and draining lymph nodes were examined for DC vaccine recovery at the injection site and at the draining lymph node one day following inoculation to evaluate the influence of neutrophils on the migratory capacities of the vaccine. The numbers of DCs in these tissues from mice that received anti-Ly6G and the DC vaccine were compared to mice that received the DC vaccine only, and negative control mice that received phosphate-buffered saline (PBS). Significant differences were determined using a one-way analysis of variance; p-value < 0.05 (*), p-value < 0.005 (**), p-value < 0.001 (***). Graphs show the means and the standard deviation with each dot representing an individual mouse.
Figure 7. Neutrophils influenced dendritic cell (DC) vaccine recovery in the footpad and draining lymph node one day post-immunization. DC vaccines were prepared ex vivo and labelled with the fluorescent dye, carboxyfluorescein succinimidyl ester (CFSE). Female C57BL/6 mice (n = 8–16 per treatment) were treated with an anti-Ly6G antibody one day prior to and on the day of DC immunization to deplete neutrophils from the mice. The footpads and draining lymph nodes were examined for DC vaccine recovery at the injection site and at the draining lymph node one day following inoculation to evaluate the influence of neutrophils on the migratory capacities of the vaccine. The numbers of DCs in these tissues from mice that received anti-Ly6G and the DC vaccine were compared to mice that received the DC vaccine only, and negative control mice that received phosphate-buffered saline (PBS). Significant differences were determined using a one-way analysis of variance; p-value < 0.05 (*), p-value < 0.005 (**), p-value < 0.001 (***). Graphs show the means and the standard deviation with each dot representing an individual mouse.
Figure 8. Depletion of neutrophils influenced the accumulation of natural killer (NK) cells in the local draining lymph node one day following dendritic cell (DC) immunization. Anti-Ly6G was administered to female C57BL/6 mice (n = 4–12 per treatment) on the day before and the day of DC immunizations to deplete neutrophils. DC vaccines were prepared ex vivo and administered into the footpads of mice. One day post-DC immunization, the local draining lymph nodes were examined by flow cytometry for NK cell populations. Statistical analysis was performed using a one-way analysis of variance; p-value < 0.05 (*), p-value < 0.001 (***), p-value < 0.0001 (****). Graphs show the means and the standard deviation with each dot representing an individual mouse.
Figure 8. Depletion of neutrophils influenced the accumulation of natural killer (NK) cells in the local draining lymph node one day following dendritic cell (DC) immunization. Anti-Ly6G was administered to female C57BL/6 mice (n = 4–12 per treatment) on the day before and the day of DC immunizations to deplete neutrophils. DC vaccines were prepared ex vivo and administered into the footpads of mice. One day post-DC immunization, the local draining lymph nodes were examined by flow cytometry for NK cell populations. Statistical analysis was performed using a one-way analysis of variance; p-value < 0.05 (*), p-value < 0.001 (***), p-value < 0.0001 (****). Graphs show the means and the standard deviation with each dot representing an individual mouse.
Figure 9. Neutrophils influenced the functionality of natural killer (NK) cells in local draining lymph nodes two days following dendritic cell (DC) immunization. Half of the female C57BL/6 mice received anti-Ly6G to deplete neutrophils the day before and the day of immunizations, and then DC vaccines or phosphate-buffered saline (PBS) were administered (n = 4 per treatment) into hind food pads. (a) Two days later, the number and proportion of NK cells in the draining popliteal lymph nodes were compared between neutrophil-depleted and non-depleted mice. (b) NK cells were stimulated using IL-2, and the production of IFNγ (left panels) and CD107a (right panels) were measured using flow cytometry. A two-way analysis of variance was used to assess statistical significance; p-value < 0.05 (*), p-value < 0.005 (**), p-value < 0.001 (***), p-value < 0.0001 (****). Graphs show the means and the standard deviation with each dot representing an individual mouse.
Figure 9. Neutrophils influenced the functionality of natural killer (NK) cells in local draining lymph nodes two days following dendritic cell (DC) immunization. Half of the female C57BL/6 mice received anti-Ly6G to deplete neutrophils the day before and the day of immunizations, and then DC vaccines or phosphate-buffered saline (PBS) were administered (n = 4 per treatment) into hind food pads. (a) Two days later, the number and proportion of NK cells in the draining popliteal lymph nodes were compared between neutrophil-depleted and non-depleted mice. (b) NK cells were stimulated using IL-2, and the production of IFNγ (left panels) and CD107a (right panels) were measured using flow cytometry. A two-way analysis of variance was used to assess statistical significance; p-value < 0.05 (*), p-value < 0.005 (**), p-value < 0.001 (***), p-value < 0.0001 (****). Graphs show the means and the standard deviation with each dot representing an individual mouse.
Figure 10. Influence of neutrophils on natural killer (NK) cell responses in the spleen one-week post-dendritic cell (DC) vaccination. Anti-Ly6G was given to half of the female C57BL/6 mice to deplete neutrophils (n = 8 per treatment). DC vaccines generated ex vivo were administered. Controls received phosphate-buffered saline (PBS). One week later, NK cell responses in the spleens were assessed. Splenocytes were stimulated with interleukin (IL)-2, and NK cell degranulation based on expression of CD107a and production of IFNγ were measured using flow cytometry. A two-way analysis of variance was used to evaluate statistical significance; p-value < 0.05 (*), p-value < 0.005 (**), p-value < 0.0001 (****). Graphs show the means and the standard deviation with each dot representing an individual mouse.
Figure 10. Influence of neutrophils on natural killer (NK) cell responses in the spleen one-week post-dendritic cell (DC) vaccination. Anti-Ly6G was given to half of the female C57BL/6 mice to deplete neutrophils (n = 8 per treatment). DC vaccines generated ex vivo were administered. Controls received phosphate-buffered saline (PBS). One week later, NK cell responses in the spleens were assessed. Splenocytes were stimulated with interleukin (IL)-2, and NK cell degranulation based on expression of CD107a and production of IFNγ were measured using flow cytometry. A two-way analysis of variance was used to evaluate statistical significance; p-value < 0.05 (*), p-value < 0.005 (**), p-value < 0.0001 (****). Graphs show the means and the standard deviation with each dot representing an individual mouse.
Figure 11. Neutrophils did not influence dendritic (DC) vaccine-mediated education of antigen-specific CD8+ T cell degranulation or production of interferon (IFNγ) and tumor necrosis factor (TNF)α. Female C57BL/6 mice (n = 12–16 per treatment) were given anti-Ly6G followed by DC vaccines. One week later, the spleens were examined for CD8+ T cell responses to stimulation with the vaccine-loaded epitopes, ovalbumin (OVA)257-264 (SIIN), which is the immunodominant CD8+ T cell epitope, and OVA323-339 (ISQ), which is the dominant CD4+ helper T cell epitope. The CD8+ T cell responses were measured by expression of CD107a as an indicator of degranulation, and production of IFNγ and TNFα using flow cytometry. The CD8+ T cell responses in the spleens of mice that received anti-Ly6G and the DC vaccine, the DC vaccine only, PBS only, and anti-Ly6G and PBS were compared using a two-way analysis of variance; p-value < 0.05 (*), p-value < 0.005 (**), p-value < 0.001 (***), p-value < 0.0001 (****). Graphs show the means and the standard deviation with each dot representing an individual mouse.
Figure 11. Neutrophils did not influence dendritic (DC) vaccine-mediated education of antigen-specific CD8+ T cell degranulation or production of interferon (IFNγ) and tumor necrosis factor (TNF)α. Female C57BL/6 mice (n = 12–16 per treatment) were given anti-Ly6G followed by DC vaccines. One week later, the spleens were examined for CD8+ T cell responses to stimulation with the vaccine-loaded epitopes, ovalbumin (OVA)257-264 (SIIN), which is the immunodominant CD8+ T cell epitope, and OVA323-339 (ISQ), which is the dominant CD4+ helper T cell epitope. The CD8+ T cell responses were measured by expression of CD107a as an indicator of degranulation, and production of IFNγ and TNFα using flow cytometry. The CD8+ T cell responses in the spleens of mice that received anti-Ly6G and the DC vaccine, the DC vaccine only, PBS only, and anti-Ly6G and PBS were compared using a two-way analysis of variance; p-value < 0.05 (*), p-value < 0.005 (**), p-value < 0.001 (***), p-value < 0.0001 (****). Graphs show the means and the standard deviation with each dot representing an individual mouse.
Comments (0)