1. IntroductionEight major cereal crops, wheat, rice, barley, rye, oats, corn, sorghum, and millet, provide two-thirds of the world’s food supply. Each of these major cereal crops is vulnerable to at least one rust disease. Rust pathogens can induce substantial damage to cereal crops, reduce yields, and cause major economic losses [
1]. Wheat stem rust poses a major threat to wheat production worldwide because outbreaks of this disease can occur over large spatial scales and spread rapidly [
2]. Wheat stem rust infections affect the stems and leaves of wheat plants. In severe cases, wheat stem rust can reduce wheat yield by 75% or result in the death of host plants [
3].Hyperparasitic fungi parasitize other plant pathogens, and both hyperparasitic fungi and their hosts impose mutual constraints on each other’s growth [
4]. Hyperparasitic fungi are widespread in natural ecosystems and commonly parasitize filamentous fungi [
5]. These fungi might provide promising alternatives to chemical fungicides for the control of fungal diseases in plants [
6]. They have thus received much research attention from botanists for their potential to be used as biocontrol agents against plant pathogens. To date, hyperparasitic fungi have been used to control a few types of rust diseases and powdery mildew [
7]. Following the colonization of host pathogens, the hyphae of hyperparasitic fungi grow toward the edge of the colony. Once the hyphae infiltrate the cell wall of the host to obtain nutrients, a parasitic relationship is established between the hyperparasitic fungus and the host pathogen, which can induce damage to the pathogen. Many hyperparasitic fungi occur naturally in plants in the form of mold colonies.Hyperparasitic fungi effectively prevent plant pathogens from infecting their hosts; these interactions between the plant pathogen and hyperparasite can inhibit the reproductive ability of the pathogenic organism [
8,
9]. The antagonism between plant and parasitic fungi and that between parasitic fungi and their hyperparasitic fungi can inhibit the growth and reproductive capacity of pathogenic organisms and thus reduce the severity of plant diseases [
8]. Rapidly growing hyperparasitic fungi might have a lower impact on the environment compared with chemical fungicides, given that they are ubiquitous in natural ecosystems. This observation has inspired much interest in the development of strategies to use these fungi as biological control agents [
9]. Many biological antagonists have been shown to work effectively against rust fungi. The application of hyperparasitic fungi is potentially an effective strategy to prevent plant diseases. A total of 30 hyperparasitic fungal genera parasitize rust fungi, including Fusarium (Fusarium spp.), Cladosporium, Scytalidium uredinicola, and Tuberculina [
10,
11]. Parasitism of the urediniospores of rust fungi has been reported in various fungal species [
12], including C. cladosporioides, Eudarluca caricis, Microdochium caricis, Microdochium nivale, Lecanicillium lecanii, and Alternaria alternata. Several Cladosporium species, such as C. uredinicola, C. cladosporioides, C. pseudocladosporioides, and C. sphaerospermum, have been reported to parasitize fungi in the order Pucciniales [
13].Scanning electron microscopy (SEM) of the abnormal urediniospores of wheat stripe rust revealed that they were parasitized by Cladosporium spp., which inactivated the urediniospores and induced structural damage [
14]. Multiple hyperparasitic fungi have been isolated from infected branches of Pinus armandii, and the defense mechanisms of fungi against Cronartium ribicola have been characterized using SEM [
15]. The hyperparasitic fungus Alternaria alternata was first reported to parasitize wheat stripe rust in 2017, and its potential utility as a biocontrol agent was demonstrated in a previous study [
5].Current methods used for the control of stem rust include the application of fungicides, the use of stem rust-resistant varieties, and preventive agricultural measures [
16]. However, the application of nonselective fungicides can lead to a disease more difficult to manage [
9]. In addition, the heavy use of fungicides can have deleterious effects on the environment, along with animals and humans [
6]. Here, the utility of hyperparasitic fungi for the control of Pgt infection is evaluated. Our findings indicate that hyperparasitic fungi could provide an effective tool for controlling destructive rust diseases of major cereal crops. The isolates of the hyperparasitic fungi in our experiment effectively inhibit the growth of Pgt and will aid future studies aimed at the development of environmentally friendly biological agents. 2. Materials and Methods 2.1. Isolation and Purification of Hyperparasitic Fungi
We were multiplying Pgt urediniospores and conducting various experiments with urediniospores in our laboratories at Shenyang Agricultural University, Shenyang. Hyperparasitic fungi were isolated from the urediniospores of Pgt, which were cultured at 22 °C and 60% relative humidity in an artificial climate chamber. The susceptible wheat cultivar Little Club was inoculated with Pgt (34C3RTGQM) and then placed in an artificial climate chamber, with 60% relative humidity, temperatures kept at 22 °C, and a 16 h/8 h light/dark photoperiod.
The white hyphae were collected from the urediniospores of Pgt-infected wheat leaves, inoculated onto potato dextrose agar (PDA) plates using an inoculation loop following 16 d of culture, and then incubated in an inverted position at 25 °C for 7–10 d in a biochemical incubator. After extracting fresh hyphae from the edge of the fungal colony, they were transferred to fresh sterile PDA medium and incubated under the same conditions to obtain a pure culture. Five replications of these experiments were performed. The purified isolates of hyperparasitic fungus were preserved on the PDA plate.
2.2. Morphological Observations of Hyperparasitic Fungi
Hyphae with vigorous growth at the edge of the isolated and purified colonies were transferred to fresh sterile PDA medium and placed in an incubator with a 12 h/12 h dark/light photoperiod at 25 °C for 7 d to make observations of the morphology of its colonies, hyphae, and conidia. Observations of hyphal and conidial morphology were performed using an Olympus BX51 microscope.
2.3. SEM of Stem Rust Fungi and Hyperparasitic Fungi
Leaves inoculated with only Pgt urediniospores were used as the CK. The hyperparasitic of isolate fungus was placed into a fungal spores suspension (105–106/mL), which was sprayed on the Pgt-infected wheat leaves. One day after Pgt inoculation, the Pgt-infected leaves were inoculated with a suspension of isolate fungus, and kept in the artificial climate chamber for growth under the same conditions. To clarify Pgt–parasite interactions during infection, the Pgt-infected wheat leaves of hyperparasitic were cut into blocks (0.2 cm × 0.2 cm) and stored in fixative for SEM. The leaves were sampled at 0, 1, 3, 5, 7, and 9 d after smearing the hyperparasitic fungal suspension.
2.4. Molecular Identification of Hyperparasitic FungiPurified hyphae (0.1 g) of hyperparasitic fungus were placed into 2.0 mL centrifuge tubes. The OMEGA (USA) HP Fungal DNA Kit was used to extract DNA from samples. A NanoDrop Microvolume Spectrophotometer (Thermo) was used to determine the concentration of DNA. Polymerase chain reaction (PCR) was then performed to amplify the internal transcribed spacer 1 (ITS1) and internal transcribed spacer 4 (ITS4) sequences (ITS1 primer: 5′-TCCGTAGGTGAACCTGCG-3′; ITS4 primer: 5′-TCCTCCGCTTATTGATATGC-3′) [
17,
18]. Electrophoresis with a 1.0% agarose gel was performed to analyze the PCR-amplified products. The thermal cycling conditions were as follows: denaturing at 94 °C for 5 min; 30 cycles (from 94 °C for 35 s, 52 °C for 60 s, to extension at 72 °C for 90 s); to extension at 72 °C for 10 min; and 4 °C for 5 min. PCR products were separated by 1% gel electrophoresis. The extracted DNA samples of the hyperparasitic fungi of Pgt were sent to Sangon Biotech Co. (Shanghai, China) for sequencing and molecular identification. 2.5. Phylogenetic AnalysisThe sequences of Simpliciium spp. were downloaded from the NCBI and aligned by ClustalW in software MEGA10 using the default parameters [
19]. The fungal isolates were clustered based on their ITS sequences using the neighbor-joining (NJ) method using MEGA10, and the branch robustness was determined using 1000 bootstrap replications [
19]. 2.6. Hyperparasitic Fungal Inhibition Experiment
The susceptible wheat (Little Club) was used for propagating Pgt urediniospores. When the first leaf had expanded after 7 days, seedlings were inoculated with 34C3RTGQM, a predominant race of Pgt in China. The collected urediniospores of Pgt race 34C3RTGQM were diluted with water to 25 mg·mL−1 and inoculated by brush. The Pgt-inoculated plants were incubated in an artificial climate chamber at 22 °C with 16 h light photoperiod. The hyperparasitic of fungus was placed into a fungal spores suspension (105–106/mL), which was sprayed on the Pgt-infected wheat leaves. Healthy wheat leaves receiving S. lanosoniveum inoculation represented control check1 (CK1). Leaves inoculated with only Pgt urediniospores were used as CK2. Three, six, and nine days after Pgt inoculation, the plants in different pots were inoculated with suspension (105~106 spores/mL) of hyperparasite, and kept in the artificial climate chamber for growth under the same conditions. Each treatment was carried out with wheat seedlings growing in three independent pots. All treatments were placed in the same artificial climate chamber, and observation of the symptoms was performed at the same time.
2.7. Urediniospore Germination Inhibition Experiment
Seedlings of wheat (Little Club) were grown in the artificial climate chamber, waiting until the first leaf had expanded after 7 days, inoculated with urediniospores of Pgt (34C3RTGQM), and incubated in an artificial climate chamber at 22 °C. Fungal suspensions (105–106 CFU/mL) were prepared in sterile water with fresh urediniospores and hyperparasitic fungal hyphae, which were extracted with a sterilized inoculating loop. The treatment group comprised a mixture of equal volumes of urediniospore suspension and hyphal suspension. In the control group, the urediniospore suspension was mixed with equal volumes of sterile water and cultured in a biochemical incubator in the dark at 22 °C for 6 h in an artificial climate chamber. Suspensions from the treatment and control groups were transferred to glass slides to perform counts of the germinated urediniospores using a light microscope. Germinating urediniospores of Pgt produce a germ tube that is at least half the length of the urediniospores. The germination rate was expressed as a percentage based on 100 randomly selected urediniospores. Experiments were performed in triplicate.
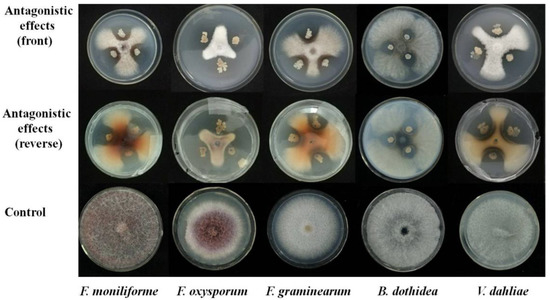
4. DiscussionStudies on the parasitism of cereal pathogenic fungi by other fungi are important for the development of biological agents for the control of diseases in economically important cereal crops. The identification of hyperparasitic fungi is critically important for understanding their biodiversity. Several hyperparasitic fungi that can parasitize plant rust pathogens have been reported in previous studies, such as Aphanocladium album, Fusarium spp., Lecanicillium spp., and Scytalidium uredinicola [
9]. Fungal parasitism has been suggested to be an effective strategy for the control of several diseases. For example, Ampelomyces quisqualis has been used to control powdery mildew on grapes and other crops [
20]. Trichoderma spp. have been used to mitigate the effects of Fusarium oxysporum on tomato plants [
21]. However, few studies have examined the efficacy of using hyperparasitic fungi for the control of Pgt on cereal crops. In our study, we isolated a fungus that could parasitize the urediniospores of Pgt. This hyperparasitic fungus could thus potentially be used for the control of Pgt, given that it inhibits the development of the urediniospores of Pgt.We studied the morphology of this hyperparasitic fungus, obtained ITS sequences, and built a phylogeny using the NJ method in MEGA10. The molecular data indicated that this fungus belonged to the genus Simplicillium, and the identification of this fungus as S. lanosoniveum had 99% bootstrap support [
22,
23].
The most practical approaches for controlling wheat stem rust that are currently used include the breeding of stem rust-resistant varieties and the application of fungicides. The loss of fungal-specific resistance in wheat varieties and the potential resistance of pathogens to fungicides are some of the major challenges in the development of biological agents for the control of wheat stem rust.

Comments (0)