1. IntroductionThe first cases of a new pulmonary disease were reported from the province of Wuhan in China in 2019. The severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) caused the disease COVID-19 and the ongoing pandemic [
1,
2]. While the majority of infected people develop only mild flu-like symptoms or are asymptomatic, the disease COVID-19 poses a serious risk of hospitalization or death to susceptible individuals [
3,
4]. So far, the most widely used measures to contain the spread of the virus have focused on the direct detection of the virus via polymerase chain reaction (PCR) or antigen-based methods and a strict quarantine regime. With a global decrease in the number of severe cases and a reduction in the number of tests used to screen contagious individuals, the serological determination of immune status will become more important in determining the risk for an individual or a population.SARS-CoV-2 consists of four major structural proteins. The spike glycoprotein (S) mediates the internalization and contains a high-affinity receptor-binding domain (RBD) for the human angiotensin-converting enzyme 2. The nucleoprotein (N) is involved in the packaging of the genome. The viral vesicular structure is formed by the membrane (M) and the envelope (E) proteins [
5,
6]. The simultaneous expression of SARS-CoV-2 structural proteins leads to the spontaneous assembly of virus-like particles (VLPs) [
7,
8]. VLPs are multi-protein structures that resemble certain viruses in their molecular composition and shape. Free of genetic material, these particles are not infectious and are therefore safe to handle.In the US, 77 serology-based tests have an active Emergency Use Authorization from the Food and Drug Administration (last updated 30 March 2022). The vast majority of these tests only measure antibodies against the spike protein or the RBD. Only 13 tests contain peptides or the full-length recombinant nucleoprotein. It has been proposed that presenting peptides from more than one protein increases the sensitivity of serological tests [
9]. However, there are only nine FDA-authorized tests that contain peptides from two proteins (N and S) and only one test that contains any peptides of the M-, N-, and S-protein. Although quite common for other viral diseases [
10,
11,
12,
13,
14], to our knowledge, there is no serological test that utilizes VLPs as antigens to detect antibodies against SARS-CoV-2.During this pandemic, SARS-CoV-2 VLPs were mostly developed as vaccine candidates [
15,
16,
17,
18] or as research tools to monitor viral entry into susceptible cells [
19,
20,
21,
22]. Surprisingly, SARS-CoV-2 VLPs have attracted very little attention in the diagnostic field.
In this proof-of-principle study, we characterized the reactivity of SARS-CoV-2 VLPs containing all four major proteins with serum from a cohort of 230 individuals. We found that high sensitivity and specificity as well as the superior performance of the VLP-based ELISA in comparison to a certified IVD validate the utility of SARS-CoV-2 VLPs as antigens to detect specific immune reactions in human serum.
2. Materials and Methods
Serum collection and ethics statement: Serum samples were obtained from different sources, such as patients of a private in-and out-patient clinic (MeoClinic, Berlin), the outpatient clinic of a physician (Dr. Omar Kamal-Eddin), from lab members and relatives, as well as in part due to the service of the Diagnostic HealthCare Solutions GmbH. Ethics approval was obtained by the ethical review board of the Charité—Universitätsmedizin Berlin (EA1/304/21) and all participants provided written consent to participate in this study. Blood was processed on the same day of withdrawal by centrifugation at 4500× g for 10 min at 4 °C. Serum samples were stored at −20 °C for short-term storage and at −80 °C for long-term storage.
Culturing of Expi293 cells: The Expi293™ Expression System Kit, composed of the Expi293 suspension adapted cell line, Expi293 transfection reagents, and Expi293 culture medium, was purchased from Thermo Fisher Scientific. The cultivation and transfection of Expi293 cells were mostly performed according to the manufacturer’s instructions. Cells were seeded at a density of between 0.3 × 106 and 0.5 × 106 cells/mL and subcultured at a concentration of 3 × 106 to 5 × 106 viable cells/mL after 3–5 days. Cell diameter, the percentage of viable cells (vitality), and the concentration of viable cells were routinely measured using the LUNA Cell Counter (Logos Biosystems, Anyang, South Korea).
Generation of SARS-CoV-2 VLPs: Virus-like particles were produced from two genetically modified Expi293 suspension-adapted cell lines. Cells were grown at 37 °C, 8% CO2 with 130–150 rpm on a Rotamax120 platform shaker (Heidolph Instruments GmbH & Co. KG, Schwabach, Germany) in Expi293 medium containing a final concentration of 100 units/mL penicillin and 100 µg/mL streptomycin. Cell line 1 (Expi_MEN), contains the human codon optimized M-(Gen Bank: QHD43419.1), E-(Gen Bank: QHD43418.1) and N-genes (Gen Bank: QHD43423.2) stably integrated into the genome. Cell line 2 (Expi_SMEN), additionally contains the S-gene (Gen Bank: QHD43416.1), including the D614G mutation and the R683A and R685A substitution, to render the furin cleavage site (FKO) non-functional, stably integrated into the genome. The induction of VLP production is controlled by the tetracycline-responsive element promoter (TRE) [
23] and can be activated by tetracycline or its analogs (e.g., doxycycline) [
24,
25]. VLP production was induced by the addition of doxycycline-hyclate (Sigma-Aldrich, Taufkirchen, Germany) at a concentration of 1 µg/mL. Approximately 96–120 h after induction, when the vitality of the cell culture was typically 40–60%, supernatants were harvested. Subsequently, cell culture supernatants were cleared by centrifugation at 2000× g for 10 min, followed by filtration with a 1.2 µm Minisart NML (Sartorius Stedim Biotech GmbH, Göttingen, Germany) followed by a 0.45 µm Millex Low Binding Durapore (PVDF) syringe filter (Merk Millipore Ltd., Tullagreen, Ireland). Clarified cell culture supernatants were continuously diafiltrated with four times the initial volume of PBS (pH 7.2) at a constant pressure of 0.124–0.165 kPa and 135 rpm using a Minimat EVO Tangential Flow Filtration System equipped with an Omega membrane with a 300 kDa cut-off (Pall Corporation, Dreieich, Germany). VLPs were precipitated from the diafiltrated retentate by the addition of PEG-it Virus Precipitation Solution (System Biosciences, Palo Alto, CA, USA) at a ratio of 1:10. The supernatants were incubated at 4 °C on a rotating shaker for 24–48 h prior to precipitation at 1500× g for 30 min. The supernatant was removed, and the VLP-containing pellet was resuspended with sterile PBS (pH 7.2) and stored at −80 °C until use.Electron microscopy: VLP-producing cells, were fixed with 2.5% glutaraldehyde (Serva, Heidelberg, Germany) in 0.1 M sodium cacodylate buffer (Serva, Heidelberg, Germany) for 30 min and postfixed with 1% osmium tetroxide (Science Services, München, Germany) and 0.8% potassium ferrocyanide II (Merck, Darmstadt, Germany) in 0.1 M cacodylate buffer for 1.5 h. Agarose-embedded samples were dehydrated in a graded ethanol series and embedded in Epon resin (Serva, Heidelberg Germany). Finally, ultrathin sections of the samples (70 nm) were stained with 4% uranyl acetate and Reynolds lead citrate [
26] (Merck). A Zeiss EM 906 electron microscope at an 80 kV acceleration voltage (Carl Zeiss, Oberkochen, Germany) equipped with a slow scan 2K CCD camera (TRS, Moorenweis, Germany) was used for image acquisition. Nanoparticle tracking analysis (NTA): NTA was applied to measure the size and concentration of VLPs in different preparations according to our previous protocol [
27]. NTA measurements were performed using a NanoSight LM10 instrument (NanoSight, Amesbury, UK) consisting of a conventional optical microscope, a high sensitivity sCMOS camera and an LM10 unit equipped with a 488 nm laser module. The samples were injected into the LM unit via the nanosight syringe pump at a constant flow rate of 50 µL/min using a 1 mL sterile syringe. Sample dilutions of 1:2000 to 1:5000 usually result in an effective particle concentration suitable for analysis with NTA (1.0 × 108 to 2.5 × 109 particles/mL). The capturing settings (shutter and gain) and analyzing settings were manually adjusted and kept constant between all samples that were recorded on the same day. NTA software (NTA 3.2 Dev Build 3.2.16) was used to capture three videos of 30 s each to analyze nanoparticle tracking data for each sample.ELISA (Enzyme-linked Immunosorbent Assay): The protocol was comparable to our previous study [
27]. S-protein (Sino Biological Inc., Beijing, China, # 40689-V08B), N-protein (charge 2020/20.7/2, 0.4 mg/mL stock, new/era/mabs GmbH, Potsdam, Germany) or SARS-CoV-2 VLPs were immobilized as antigens on Nunc Polysorb 96-well microtiter plates in 100 µL carbonate buffer per well overnight at 4 °C. Free binding sites were blocked with 250 µL per well of PBS containing 1% bovine serum albumin (BSA) for 45 min at room temperature (RT). After washing with PBS, rabbit monoclonal antibodies against spike- (1:2000, 40689-V08B, Sino Biological Inc., Beijing, China) and nucleoprotein (1:5000, 40143-R019, Sino Biological Inc., Beijing, China) or human serum (1:50–1:100) were incubated with the immobilized antigen in 100 µL PBS containing 0.05% Tween20 (PBS-T) and 1% BSA at RT for 1 h. A detergent-containing buffer was chosen to facilitate access of antibodies to epitopes inside the lumen of the VLPs. After washing, plates were incubated at RT with a horseradish peroxidase (HRP)-conjugated goat anti-rabbit or goat anti-human IgG antibody (Dianova GmbH, Hamburg, Germany) diluted 1:10,000 in PBS-T containing 1% BSA. Unbound conjugated molecules were removed by washing with PBS. The colorimetric reaction using 100 µL tetramethylbenzidine (TMB, Carl Roth GmbH, Karlsruhe, Germany) as a substrate was stopped with 100 µL of 0.125 M H2SO4 per well after 10–30 min. Absorbance was immediately measured with an Epoch microplate reader at λ = 450 nm and subtracted by the absorbance of the reference wavelength at λ = 620 nm. For the quantitative approximation of N- and S-protein concentrations in VLP samples, the linear range of a 2-log dilution series (triplicate) of the respective proteins was taken as a reference.Western blot: Samples were prepared according to the NuPAGE Technical Guide of Invitrogen. After denaturation in NuPAGE LDS sample buffer with DTT 50 mM for 10 min at 70 °C, the samples and markers were run on a Novex bis-tris gradient gel (4–12%, Thermo Fischer Scientific) using NuPAGE MOPS SDS running buffer and subsequently blotted on a Novex 0.45 µm nitrocellulose membrane (LC2001, Thermo Fischer Scientific). The membranes were blocked for 1 h in PBS containing 1% BSA, and subsequently incubated with a rabbit-anti-nucleoprotein primary antibody (1:5000, 40143-R019, Sino Biological Inc, Beijing, China) or human serum in PBS containing 1% BSA and 0.2% PBS-T at 4 °C overnight on a shaker. Human serum from a double-vaccinated individual (AstraZeneca) was used to detect the S-protein alone or S-protein and N-protein, respectively. High levels of anti-S IgGs were confirmed in the respective human sera by ELISA. HRP-coupled goat anti-human or goat anti-rabbit (Dianova GmbH, Hamburg, Germany) secondary antibodies were incubated at a dilution of 1:10,000 in PBS-T for 3 h at RT. Proteins were detected by chemiluminescence using respective kits from Biozym Scientific GmbH (Hessisch Oldendorf, Germany) and submitted for image analysis with ImageJ [
28].
Data evaluation and statistical analysis: Statistical analysis was performed with Prism 8 (GraphPad, San Diego, CA, USA). p values of less than 0.05 were considered statistically significant. Differences between the groups were tested with ANOVA or respective nonparametric methods (Kruskal–Wallis test), followed by multiple comparison (Dunnett’s or Dunn’s tests). Significance levels were marked by asterisks, where * corresponds to p < 0.05, ** p < 0.01, *** p < 0.001 and **** p < 0.0001.
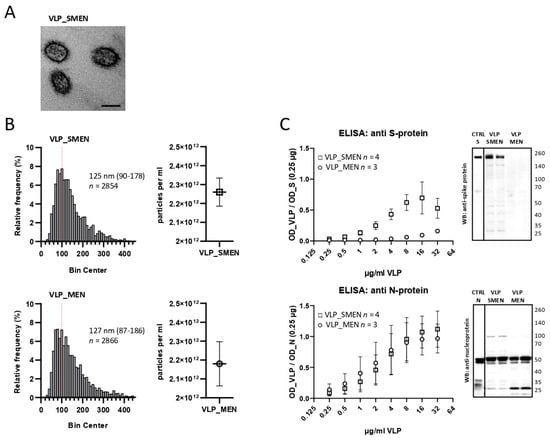
4. DiscussionThis is the first report using virus-like particles in a solid-phase immunoassay to detect antibodies against SARS-CoV-2. In contrast, VLPs are used in some vaccination approaches [
18,
29]. We were able to generate stable particles (127 nm for MEN VLP, 125 nm for SMEN VLP) of an expected size similar to the natural virus (60 to 140 nm as reported by [
2]). The appropriate virus-like assembly was illustrated by electron microscopy. ELISA and Western blot experiments validated the presence of the N-and S-protein. The presence of the M- and E-protein can be implied because they are essential for the formation of particles [
7,
8,
30,
31].
Theoretically, there are some advantages to using VLPs instead of isolated single proteins or natural virus isolates to produce an antigen-coated surface for solid-phase immunoassays to detect anti-virus antibodies: (i) There is a greater likelihood that the antigens will retain their natural conformation. (ii) By utilizing the third dimension, there could be a greater amount of overall antigens available. (iii) Due to the lack of genetic material, VLPs are not infectious. (iv) When VLPs contain all of the virus proteins, some additional epitopes might be available. Therefore, an ELISA using VLPs may be able to detect more antibodies in a serum sample, resulting in a higher sensitivity.
A similar VLP-based diagnostic ELISA was developed for the detection of human papillomavirus type 16, which is a risk factor for cervical cancer [
10,
12,
32,
33]. VLP-based ELISAs are frequently used in veterinary medicine to detect antibodies against different viruses such as swine vesicular disease virus [
11], porcine circovirus type 2 [
13], or Senecavirus A [
14]. We were surprised that there is no VLP-based ELISA for the detection of antibodies against SARS-CoV-2 available today.
The VLP-ELISA that was developed in this study performed notably well. The small clinical study clearly demonstrated that the diagnostic performance met our expectations: 100% correct results for negative samples but higher sensitivity for positive samples compared to a commercially available CE-marked S-protein S1-subunit IVD kit (late infection 97.7% versus 95.5%, vaccinated 98.5% versus 84.2%). The sensitivity in the early stage (less than 3 weeks) of infection was remarkable: 35.3% versus 0.0%. It is important to minimize the diagnostic gap between the time of infection and the time of a positive antibody test. When considering the sensitivity and the specificity, the overall performance was better when using VLPs as an antigen rather than the individual N- or full-length S-protein alone.
Here we present a proof-of-concept for a VLP-based ELISA in SARS-CoV-2 infection. The preliminary clinical data show very high sensitivity and specificity. These data should be confirmed by a larger clinical study.







Comments (0)