
Remember me
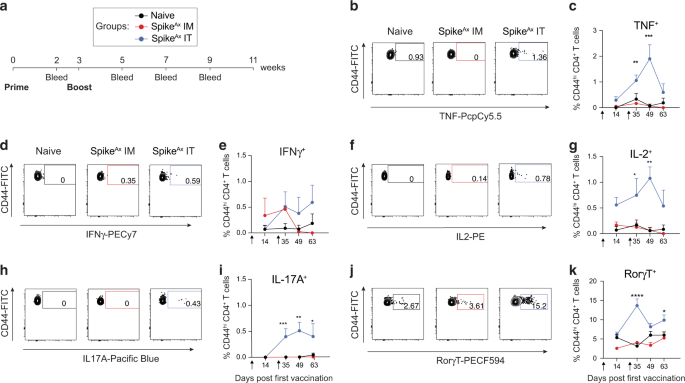


To investigate the development of allergen-specific antibodies and murine food allergy, mice were sensitized to hen’s egg by intra-tracheal treatment with egg white (EW)/egg yolk plasma (EYP) followed by repeated oral gavage (o.g.) with EW and EYP39. Starting at week four, mice were examined for diarrhea and hypothermia in response to o.g. egg inoculation (Fig. 1a). Depending on the individual response, mice could be assigned into three different groups characterized by the development of either diarrhea alone, diarrhea, and hypothermia, or neither (Fig. 1b). The development of diarrhea and hypothermia was strongly associated with increased EW-specific IgE and IgG1 titers (Fig. 1c). IgA and IgG2a anti-EW titers were also highest for mice that had both diarrhea and hypothermia at challenge 13 and 15, respectively, but not after challenge 7, while IgG2b anti-EW titers were highest for mice that had both diarrhea and hypothermia after challenge 7, but not challenge 15. Consistent with a protective effect of IgG138, allergen-specific IgG1 to IgE ratios averaged more than tenfold higher in mice with neither disease feature as compared to mice that had one or both features after 13 challenges.
Fig. 1: The food allergy model.
Mice were sensitized to and challenged with EG/EYP and analyzed for the development of allergic symptoms. a Experimental outline. b Based on the development of symptoms, mice were categorized into different groups: mice that had at least 2× diarrhea but never a temperature drop ≥ 1 °C in response to a challenge (blue dot, diarrhea only; n = 11); mice that had at least 1× diarrhea and 2× a maximum temperature drop (max. ΔT) ≥ 1 °C in response to a challenge (red triangle, both symptoms; n = 14) and mice that never showed diarrhea or a max. ΔT ≥ 0.5 °C (black triangle No symptoms; n = 28), as indicated. The average drop in body temperature (left) and the occurrence of diarrhea (right) are shown. Data presented as mean ± SEM. c Mice were bled and EW-specific antibodies were measured by ELISA. EW-specific Ig subclasses and the ratio of IgG1 to IgE are shown. Pooled serum from EW/EYP-allergic mice was used as a standard that was defined as having 100 units of each Ab. Each symbol represents data form one mouse. Total number of mice: 53. No sample was excluded from the analysis. Statistics: two-way ANOVA with Bonferroni post test; not significant (no mark): p > 0.05 *p < 0.05, **p < 0.01, ***p < 0.001. Mean ± SEM is shown.
Follicular and extrafollicular responses contribute to IgE productionIn order to investigate the IgE and IgG1 repertoires, samples were taken from the intestine-draining mesenteric lymph nodes (mLN) and bone marrow (BM) of two naive, and five allergic mice that had developed a temperature drop ≥ 1 °C and diarrhea after seven weeks of repeated allergen challenges (13 o.g. egg inoculations). cDNA was generated from the individual samples, barcoded, and amplified, after which the BCR-repertoires encoding the antigen-binding VDJ-IgH-regions that were 5′ to Cε or Cγ1 sequences were analyzed by NGS (Fig. S1).
Between one and four hundred thousand sequences were analyzed for each sample (Table S1). IgG1 sequences were approximately four- and nineteen-times more abundant than IgE sequences (Fig. 2a). Individual BCR clonotypes were defined as sharing a unique VDJ rearrangement with conserved CDR3-IMGT anchors (cysteine C 104, tryptophan W 118 or phenylalanine F 118). IgE sequences and IgE clonotypes were approximately 30% less frequent in BM than in mLN. The ratios of IgG1/IgE sequences and of IgG1/IgE clonotypes were 5-8-fold higher in BM compared to mLN (Fig. 2b), indicating that the accumulation of IgE+ plasma cells within the BM is more tightly restricted than the accumulation of IgG1+ plasma cells in this tissue. This conclusion is consistent with results suggesting that IgE and IgG BCRs differently impact B-cell fate and the development of long-lived plasma cells40,41 and that IgG+ plasma cells have a better BM homing ability than IgE+ plasma cells42. While the majority of IgE and IgG1 clonotypes showed VDJ-hypermutation, VDJ-hypermutation was absent in approximately 7–12 % of IgE and IgG1 in mLN and BM clonotypes (Fig. 2c). Approximately half of these unmutated clonotypes were clonally expanded, with more than 50 copies per clones (Fig. 2d).
Fig. 2: IgE and IgG1 sequences and clonotypes.
cDNA from naive mice and allergic mice challenged o.g. 13 times with EG/EYP was analyzed by NGS. Individual clonotypes within the IgG1 and IgE IgH-repertoires were defined by unique VDJ-rearrangements. a Number of sequences and clonotypes in BM and mLN, as indicated. Colored bars represent the mean. b Ratios of IgG1 to IgE sequences and ratios of IgG1 to IgE clonotypes, as indicated. c Number and percentages of hypermutated and unmutated IgE and IgG1 clonotypes. Representative data from two samples from BM and mLN each are shown. d Clone size distribution of unmutated IgE clonotypes. Representative data from mLN and BM of one mouse are shown, as indicated. e Upper panel: clone size distribution of all mutated IgE clonotypes. Lower panel: Clone size distribution of mutated IgE clonotypes with a clone size of 10 or less. f Mean copy numbers. Each symbol represents the average copy numbers of mutated IgG1 clonotypes (muIgG1), mutated IgE clonotypes (muIgE), unmutated IgG1 clonotypes (umIgG1) and unmutated IgE clonotypes (umIgE) from the mLN or BM of one mouse, as indicated. g Number of high copy number clones, defined as clones with copy numbers greater than the average umIgE or umIgG1 copy number, respectively. a/b: samples from 5 allergic mice and 2 naive mice from one experiment were analyzed. One BM sample was excluded due to insufficient cDNA quality (mLN: n = 7, BM: n = 6). c–f samples from mLN of 5 allergic mice from one experiment and pooled samples from BM of 7 allergic mice from two experiments are shown (mLN: n = 5, BM: n = 7). No sample was excluded from the analysis. Total number of mice: 7. Statistics a-c: Mann Whitney’s nonparametric test for comparisons within the BM or mLN; Wilcoxon nonparametric matched paired test for comparisons between BM and mLN. Data presented as mean ± SEM. Statistics d–g: pairwise t-test. *p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001; ****p ≤ 0.0001.
Mutated IgE clones were more abundant, but their clone size was much more variable. While a few mutated IgE clones had a huge clone size with copy numbers from 100 to >1000, the great majority of mutated IgE clones was present in copy numbers below 10 (Fig. 2e). Comparable numbers of unmutated and mutated IgE clonotypes were present for clonotypes present in copy numbers >50.
As a result of the differences in clone size distribution, unmutated IgE and IgG1 clonotypes had higher mean copy numbers than mutated clones (Fig. 2f), but clonotypes with high copy numbers (copy numbers ≥ the mean for unmutated IgE and IgG1) were more abundant among the mutated clonotypes (Fig. 2g).
These results are in accord with the co-existence of a follicular response and an ongoing recruitment of naive B cells into an extrafollicular response, with the former yielding hypermutated and highly selected IgE clonotypes and the latter forming unmutated IgE clonotypes that underwent relatively little clonal selection. This finding is consistent with a recent study indicating that an extrafollicular response contributes to chronic IgE production in patients suffering from allergic rhinitis43.
BCR clonotypes vary in IgE to IgG1 ratiosThe great majority of individual mutated and unmutated IgE sequences were also present as IgG1 sequences (Table 1). This is consistent with a close relationship between the IgE and IgG1 repertoires and the development of most secondary IgE responses from the IgG1 memory compartment24,29,44.
As expected from the overall excess of IgG1 compared to IgE sequences, most shared clonotypes showed a several-fold excess of IgG1 compared to IgE copies. However, other clones shared between IgE and IgG1 showed a strong bias towards IgE, containing 5 to 10-fold more IgE than IgG1 copies (Fig. 3a, b). Based on these data, clones found within the IgE repertoire were classified as IgE-biased (IgEE, with an IgE/IgG1 copy number ratio ≥ 2) or IgG1-biased (IgEG, with an IgG1/IgE copy number ratio ≥ 2).
Fig. 3: IgEG and IgEE clonotypes.
Sequences from allergic mice were analyzed after 13 o.g. allergen challenges. a Shared clonotypes (VH-genes) were ordered in accordance to the ratios of their IgE to IgG1 copy numbers. Representative data from one mLN and one BM sample are shown. b Shared clonotypes from the 50 most abundant clonotypes. c Shared clones were classified as IgE-biased (IgEE, IgE/IgG copy number ratio of ≥ 2), or IgG1-biased (IgEG, IgG/IgE copy number ratio of ≥ 2). Copy numbers of mutated IgEE (mIgEE), mutated IgEG (mIgEG), unmutated IgEE (umIgEE), and unmutated IgEG (umIgEG) from BM and mLN are shown, as indicated. Each symbol represents the mean copy numbers form one mouse for the IgE subtype indicated. mLN samples from 5 allergic mice from one experiment and BM samples from 7 mice pooled from two independent experiments were analyzed (mLN: n = 5, BM: n = 7). No sample was excluded from the analysis. d Hypermutation rates of IgEG and IgEE. Mean percentage of non-silent mutations within the IgH-CDR3 regions of mutated IgEE (muIgEE), mutated IgEG (muIgEG), unmutated IgEE (umIgEE) and unmutated IgEG (umIgEG) are shown for BM and mLN, as indicated. Data presented as mean ± SEM. Statistics: pairwise t-test. *p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001; ****p ≤ 0.0001.
As indicated by their copy numbers (clone size), IgEE and IgEG showed different levels of clonal expansion (Fig. 3c). Mutated IgEE clone sizes averaged > 2-fold larger than mutated IgEG clones in both mLN and BM. Clone sizes of unmutated IgE subtypes did not show significant differences. Consistent with the idea that the increased clonal expansion of mutated IgEE compared to IgEG is the consequence of differential selection within the germinal center, the two mutated IgE subtypes showed different hypermutation rates in the CDR regions (Fig. 3d).
In order to investigate whether IgEE and IgEG clones represent a unique feature of our allergy model, the BCR repertoires of the IgE and IgG1 compartments in BM of two naive mice were analyzed. The number of distinct clonotypes was low, as expected for unimmunized mice (Fig. S2). The few individual clonotypes and the high degree of variation is consistent with the undefined immune status and overall low-level immune stimulation of naive mice. The detectable IgE clonotypes showed also a bias for either IgE or IgG1. Next, we re-analyzed published sequences from mice infected with Nippostrongylus brasiliensis31, a helminth parasite that induces a strong IgE response. IgEE and IgEG were also present in the N. brasiliensis-infected mice, and within the infection model there were also mutated and unmutated IgE subtypes found (Fig. S2b, Table S2).
In our murine egg-allergy model, approximately 10–20% of IgE clonotypes were unique, i.e. not detectable as IgG1 clonotypes. But these unique clonotypes were present in comparably low copy numbers (Fig. S3).
Together, these data indicate that IgEE and IgEG clonotypes represent a common phenomenon, seen in naive, allergic and helminths infected mice. They show that the relative IgE to IgG1 production is controlled at the level of individual B-cell clones, with mutated IgEE and IgEG subjected to differential hypermutation and clonal selection. Hence, individual BCRs themselves seem to have a strong impact on the nature of the IgE response and its relationship to the IgG1 response.
BCR properties affect the formation of extrafollicular and follicular IgEE and IgEGNet charge, aliphatic index, hydrophobicity, and isoelectric points are important parameters that determine the antigen-binding priorities and polyreactivity of individual BCRs45. These physiochemical properties were calculated using the “Peptide” package in the R program. Overall, mutated IgEE and IgEG clonotypes exhibited similar physiochemical properties, indicating that they are not relevant for the generation of individual IgE to IgG1 ratios. However, the mutated IgEE and IgEG clonotypes differed in the physiochemical properties of their unmutated counterparts (Fig. 4). In the BM, mutated and unmutated IgEG significantly differed in their average net charges. In the mLN, mutated and unmutated IgEG exhibited different aliphatic indexes and showed considerably different hydophobicities, and mutated and unmutated IgEE exhibited different isoelectric points. The observed greater importance of antigen-binding properties in the BM compared to mLN are consistent with the idea that the former contains highly selected clonotypes.
Fig. 4: Physiochemical properties of IgEG and IgEE.
Sequences from mLN and BM of allergic mice were analyzed after 13 o.g. allergen challenges. Physiochemical properties of IgEE and IgEG clones were calculated using the “Peptide” package in the R program. Aliphatic index, hydrophobicity index according to Kyte-Doolittle, net charges and isoelectric point (PI), are shown for mutated IgEE (muIgEE), mutated IgEG (mIgEG), unmutated IgEE (umIgEE) and unmutated IgEG (umIgEG). mLN samples from 5 allergic mice from one experiment and BM samples from 7 mice pooled from two independent experiments were analyzed. Two BM samples did not contain detectable umIgEE/ umIgEG. a Data from BM (n = 5–7). b Data from mLN (n = 5). Each symbol represents the respective average ratios form one mouse. Data presented as mean ± SEM. Statistics: pairwise t-test. *p ≤ 0.05; **p ≤ 0.01.
These findings indicate that the differential binding of individual BCRs to antigens having distinct physiochemical properties has an impact on hypermutation and clonal selection, but does not determine the development of IgEE versus IgEG.
BCR signaling strength and IL-21 affect the ratio of IgE to IgG1 productionNext, we tested the possibility that differential BCR-signaling strength, which may mimic higher or lower affinities/avidities of BCRs for their cognate antigen, has an impact on the development of IgE versus IgG1. Therefore, we used a modified version of an advanced culture system that allows in vitro generation of germinal-center-like cells46. Naive B cells or sorted IgG1+ B cells were isolated from spleens of naive mice and stimulated with IL-4 and varying amounts of anti-IgM or anti-Ig kappa F(ab′)2 fragments. Anti-Ig kappa Ab stimulates both the IgM and IgD BCRs, which seem to be relevant for BCR activation by certain types of antigen7; and can also stimulate IgG1+ B cells.
Both anti-IgM and anti-Ig kappa F(ab’)2 induced a dose-dependent calcium flux (Fig. 5), indicating that titrated concentrations of these reagents can mimic the differential signal strength that would characterize Ag stimulation of low or high-affinity BCRs. At day 4 of culture the frequencies of IgE+ and IgG1+ cells were analyzed by flow cytometry (Fig. 6a). With increasing BCR stimulation, the frequencies of IgE+ and IgG1+ cells decreased (Fig. 6b). The finding that IgG1+ and IgE+ cells were reduced is in accordance with the finding that BCR signaling can inhibit class switch to these Ig classes47. However, the decrease was greater for IgE+ cells and the ratio between IgE+ and IgG1+ cells dropped from approximately 0.14 in the absence of BCR stimulation, to below 0.03 at the highest concentration of stimulating antibodies. Therefore, we speculated that BCR stimulation could inhibit not only direct class switching from IgM to either IgG1 or IgE, but also the ability of IgM+ cells to switch sequentially to IgE via an IgG1+ intermediate, which could explain why IgE+ cells decreased more rapidly than IgG1+ cells. In line with this idea, sequential class switch-specific switch circles were reduced upon BCR stimulation (Fig. S4). Of note, switch circles for direct class switch were hardly detectable. This finding may indicate that in our cultures, sequential class switch might be the dominant mechanism of IgE formation. This is in line with earlier findings in B-cell cultures stimulated with LPS + IL-448.
Fig. 5: BCR crosslinking induced calcium flux.
Single-cell suspensions from spleen were stained for IgM and IgD. After incubation with CalbryteTM 520 AM, separate samples were treated with various concentrations of polyclonal anti-IgM, or anti-Ig-kappa F(ab′)2, as indicated. Calcium flux was measured by flow cytometry (LSRII, BD, using a Flow Jo 10.7.1 software). a Representative histogram plots for IgMhigh/IgDlow naive B cells and IgMhigh/IgDlow marginal zone B cells, as indicated. b Statistical analysis of calcium flux after incubation with increasing concentrations of polyclonal anti-IgM F(ab′)2, for IgMhigh/IgDlow naive B cells, IgMhigh/IgDlow marginal zone B cells and IgMneg/IgDneg cells. Each symbol represents one cell culture well (n = 5 for all conditions). Representative data from one of three independent experiments, median and range are indicated. No sample was excluded from the analysis. Statistics: Friedmann-Test was calculated using R. ***p ≤ 0.001.
Fig. 6: BCR stimulation alters the IgE to IgG1 ratio for class-switched cells.
Naive B cells were cultured together with CD40L / BAFF-transfected feeder cells and IL-4, and stimulated with various concentrations of anti-IgM F(ab’)2 fragments. Subsequently, the frequencies of IgE+ and IgG1+ cells were analyzed by flow cytometry. Dead cells, debris, doublets, and feeder cells were excluded by Life/Dead stain and forward/sideward scatter and B cells were identified by CD19 expression. a Representative FACS plots of CD19+ B cells on day four of culture, stimulated with increasing concentrations of anti-IgM, as indicated. b Percentages of IgE+ and IgG1+ B cells, and the ratios between IgE:IgG1+ cells are shown at day four of culture, as indicated. c IgG1+ cells were sorted from B-cell cultures by FACS-sorting, stimulated with IL-4 and various concentrations of anti-Ig kappa F(ab’)2 fragments, re-cultured for another three days and subsequently analyzed by flow cytometry. Percentages of IgE+ and IgG1+ cells in cultures stimulated with various concentrations of anti-Ig kappa F(ab′)2 fragments, as indicated. Statistics: Each dot represents data from a culture of cells from one mouse (n = 8 for all conditions using anti-IgM; (n = 6 for all conditions using anti-Ig kappa). No sample was excluded from the analysis. b Data are pooled from three independent experiments. Data presented as mean ± SEM. Statistics: Friedman-test, ***p ≤ 0.001. C: paired t-test, **p ≤ 0.01; ****p ≤ 0.0001.
In order to further address the idea that BCR signaling inhibits sequential class switching, IgG1+ cells were sorted from cultures and re-cultured in the presence of IL-4 and increasing amounts of anti-kappa Ig F(ab’)2. After one day, without BCR stimulation, between 12 and 22 % of the isolated IgG1+ cells had switched to IgE and lost IgG1 expression. Increasing BCR stimulation inhibited the appearance of IgE+ cells in a dose-dependent manner but increased the percent of IgG1+ cells back to nearly 100 % (Fig. 6c).
These results extend the finding that BCR signaling inhibits class switch to IgG1 and IgE47 by showing that increasing BCR signaling also effects sequential class switching of IgG1+ cells to IgE, eventually reducing the ratio of IgE/IgG1+ cells.
Antigen-binding affinities of individual BCRs also affect antigen-uptake49,50, which could modulate the interaction of antigen-specific B cells with T follicular helper cells which provide IL-21. Therefore, we tested the effects of IL-21 in our culture system. IL-21 promoted the formation of IgG1+ cells, while inhibiting IgE, eventually shifting the ratios of IgE+ to IgG1+ cells from approximately 0.1 to above 0.2 (Fig. 7a, b).
Fig. 7: IL-21 reduces the IgE to IgG1 ratio for class-switched cells.
Naive B cells were cultured together with CD40L / BAFF-transfected feeder cells and IL-4. At day 0 and day 1, various concentrations of IL-21 were added. The frequencies of IgE+ and IgG1+ cells were analyzed by flow cytometry at day 4. Dead cells, debris, doublets and feeder cells were excluded by Life/Dead stain and forward/sideward scatter and B cells were identified by CD19 expression. a Representative FACS plots of CD19+ B cells, stimulated with IL-21 in concentrations of 0, 10 or 20 ng/ml, as indicated. b Statistical analysis of the percentages of IgE+ and IgG1+ B cells, and the ratios between IgE:IgG1+ cells. Representative data from one of two independent experiments are shown. No sample was excluded from the analysis. Data presented as mean ± SEM. Statistics: Each symbol represents data from a culture of cells from one mouse (n = 5 for all conditions). Unpaired parametric t-test *p ≤ 0.05; **p ≤ 0.01; ****p ≤ 0.0001.
Together, these data suggest that BCRs could affect the ratios of individual B-cell clonotypes through both the strength of the BCR signaling, and via modulation of B-cell interaction with T follicular helper cells providing IL-21, at least in our murine models.
IgG1+ cells with both low and high hypermutation rates can give rise to IgEIn order to determine whether IgG1+ cells that had undergone extensive GC reactions could still undergo class switch to IgE, we analyzed the clonal distribution of hypermutation rates and mutational trees in our food allergy model. Most shared IgE clones showed low to moderate hypermutation rates below 12 mutations, less than 10% showed up to 27 mutations (Fig. 8a, b). Because highly hypermutated IgG1 clones cannot give rise to moderately mutated IgE clones, these results indicate that low to moderately hypermutated IgG1 clones contribute most of the sequential class-switched IgE.
Fig. 8: Clonal distribution of somatic hypermutation rates and lineage tree analysis.
cDNA from allergic mice challenged o.g. 13 times with EG/EYP was analyzed by NGS. Individual clonotypes within the IgG1 and IgE IgH-repertoires were defined by unique VDJ-rearrangements. a Distribution of somatic hypermutation (SHM) rates of shared IgE and IgG1 clonotypes in one representative samples. b Statistical analysis of the distribution of somatic hypermutation of shared clonotypes in samples from BM of a total of 7 mice from two independent experiments (n = 7). Number of shared IgE clones with various hypermutation rates are shown, as indicated. c Analysis of lineages trees of highly mutated BM IgE clonotypes. Three samples containing less than 10 highly mutated IgE clonotypes were excluded from the analysis (n = 4). The number of highly mutated IgE (≥ 12 mutations) clonally related to highly mutated IgG1 (≥ 12 mutations) and low/moderately mutated IgG1 (≤ 11 mutations) are shown. (b, c) Each symbol represents data from one mouse and colored bars represent the mean. In total, lineage trees of 40 highly mutated IgE clonotypes were analyzed. d–h Examples of lineage trees of highly mutated IgE clonotypes are shown. Data presented as mean ± SEM. Statistics: Paired parametric t-test *p ≤ 0.05; **p ≤ 0.01.
However, lineage tree analysis indicated that approximately 80% of highly mutated IgE clonotypes (12 or more mutations) were related to highly mutated IgG1 clonotypes (Fig. 8c–h). Hence, class switch to IgE preferentially occurs before B cells have passed through extensive germinal center reactions, but highly mutated IgG1 clonotypes are still able to switch to IgE. The low proportion of highly mutated IgE derived from IgG1 progenitors with fewer mutations also indicates that once switched to IgE, the cells tend to avoid participating in further germinal center reactions.
Together, our findings indicate that the physiochemical properties of individual BCRs affect their mutation rates and clonal selection and that the BCR affects the ratio of IgG1 to IgE formation through its individual antigen-signaling strength and soliciting T-cell help. This mechanism could bias high-affinity B cells, receiving strong antigen-induced BCR signaling and IL-21 from T follicular helper cells, to favor IgG1 over IgE production.
Comments (0)