
Remember me
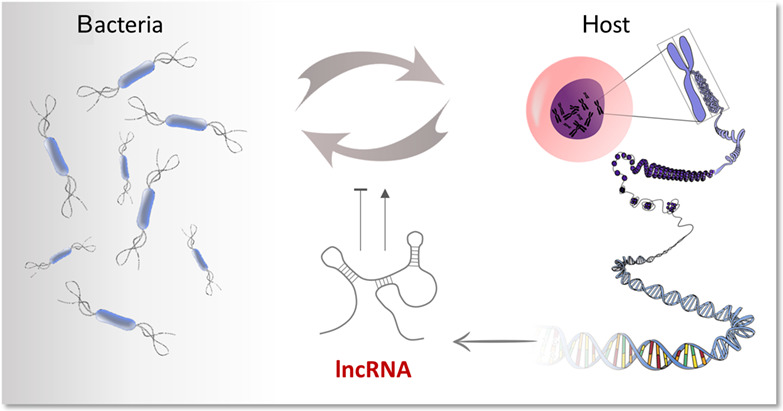
The human skin and mucosal surfaces closely interact with bacterial communities, which colonize the host as symbionts and contribute to homeostasis. During bacteria–host co-evolution several microbial species have developed strategies to overcome these barriers and manifest an infection, for example, by altering phagosomal maturation, cell death and immune response pathways (Bengoechea & Sa Pessoa, 2019; Rocco & Irani, 2011; Sindhwani & Arya, 2017). By combating such facultative or obligate pathogenic bacteria, the cellular immune system plays a key role in the maintenance of organismal homeostasis. Whereas the innate immune system confers immediate defense against invading pathogens, long lasting protection is provided by the adaptive immune system.
Different from adaptive immune cells, which sense foreign antigens by a nearly infinite arsenal of B- and T-cell receptors (BCRs, TCRs), generated by somatic DNA recombination, the innate immune system relies on germ-line encoded pattern recognition receptors (PRRs). Several PRRs are deeply conserved throughout bilaterian animal evolution and sense equally conserved pathogen structures, referred to as pathogen associated molecular patterns (PAMPs) (Akira et al., 2006), as well as damage associated molecular patterns (DAMPs), released by damaged tissue and necrotic cells (Rathinam & Fitzgerald, 2016; Takeuchi & Akira, 2010). A prototypical example is the plasma membrane standing Toll-like receptor 4 (TLR4), a member of a PRR family originally identified in fruit flies (Lemaitre et al., 1996). Mammalian TLR4 senses extracellular bacterial cell wall lipopolysaccharide (LPS) and activates two distinct intracellular signaling programs (Figure 1). Upon activation of TLR4, the intracellular adapter Myeloid Differentiation Primary Response Protein 88 (MyD88) forms a complex with Interleukin-1 Receptor Associated Kinase 1 (IRAK1) (Lin et al., 2010). This results in auto-phosphorylation of IRAK1 and its dissociation from MyD88 (Jiang et al., 2002). IRAK1 subsequently promotes TNF Receptor Associated Factor 6 (TRAF6) auto-ubiquitination and Transforming Growth Factor-β-activated Kinase 1 (TAK1) ubiquitination. TAK1 in turn forms a complex with the TGF-Beta Activated Kinase 1 Binding Proteins TAB1, TAB2, and TAB3 (Z. J. Chen, 2012) to trigger C-Jun N-Terminal Kinase (JNK)/p38 Mitogen-Activated Protein Kinase (MAPK) dependent activation of the transcription factor Activator Protein 1 (AP1) or IKK complex dependent activation of the Nuclear Factor Kappa B (NFκB) transcription factor (Akira et al., 2006; Takeuchi & Akira, 2010). Upon binding of TAK1 to the IKK complex via ubiquitin chains, Inhibitor Of NFκB Kinase Beta (IKKβ) is phosphorylated and activated. This triggers phosphorylation, ubiquitination and subsequent degradation of the cytoplasmic NFκB inhibitor IκBα, resulting in nuclear translocation of NFκB and activation of the majority of LPS-induced pro-inflammatory immune genes (Akira et al., 2006; Takeuchi & Akira, 2010) (Figure 1). In addition to MyD88, TLR4 can also signal through the TIR Domain-Containing Adapter Protein Inducing IFN-Beta (TRIF), which interacts with TNF Receptor Associated Factor 3 (TRAF3). Upon auto-ubiquitination, TRAF3 promotes phosphorylation of TANK Binding Kinase 1 (TBK1), which in turn phosphorylates the Interferon Regulatory Factor 3 (IRF3) transcription factor (Akira et al., 2006; Takeuchi & Akira, 2010). This final step of the TIRF-dependent signaling arm is supported by the TBK1 platform protein and ubiquitin-reader optineurin (OPTN) (Gleason et al., 2011). Upon nuclear translocation, IRF3 induces transcription of type I interferon (IFN) genes (Akira et al., 2006). Upon release, type I IFN binding to IFN alpha receptor (IFNAR) dimers triggers Janus Kinase (JAK) transphosphorylation and JAK-dependent Signal Transducer And Activator Of Transcription (STAT) phosphorylation (Rawlings et al., 2004) (Figure 1). Activated STAT proteins, such as STAT1, promote the transcription of interferon-stimulated genes (ISGs) in a wide range of target cell types (Figure 1) (Swiecki & Colonna, 2011). Proteins encoded by ISGs in turn induce an infection-refractory state, for example, through inactivation of viral transcription and translation pathways (Kagan & Barton, 2014; Schneider et al., 2014; Takeuchi & Akira, 2010; Trinchieri, 2010). Besides interference with viral replication, type I IFNs are increasingly recognized to promote defense against bacterial pathogens (Boxx & Cheng, 2016). Type I IFN responses, can, however, also contribute to infection-induced pathologies and for instance exacerbate pulmonary tuberculosis or increase susceptibility to Listeria monocytogenes infection (Stifter & Feng, 2015). TLR4 is the only PRR capable of inducing both the MyD88- and the TRIF-dependent immune signaling pathway (Takeuchi & Akira, 2010). The human TLRs signaling through MyD88 are the flagellin sensor TLR5, the single-stranded RNA (ssRNA) sensors TLR7 and 8, the CpG DNA receptor TLR9 and the lipoprotein sensor TLR2, which acts in conjunction with TLR1 or 6. TLR3, which senses double-stranded RNA (dsRNA), signals through the TRIF pathway (Takeuchi & Akira, 2010). Other PRR families are the C-type lectin receptors (CLRs), the RigI-like receptors (RLRs) and the Nucleotide-binding Oligomerization Domain (NOD)-like receptor (NLR) family. While the CLRs primarily participate in antifungal defense, RLRs sense viral and bacterial nucleic acids and induce type I IFN expression through the Mitochondrial Antiviral Signaling Protein (MAVS), instead of the TRIF signaling adapter (Takeuchi & Akira, 2010). The NLR family members NOD1 and NOD2 sense intracellular bacterial peptidoglycans and activate NFκB and AP1 through a signaling cascade depending on Receptor Interacting Serine/Threonine Kinase 2 (RIP2) instead of MyD88 (Takeuchi & Akira, 2010). Other members of the NLR family are engaged in the inflammasome pathways, which are activated by cytosolic DAMPs and PAMPs. While NLR Family Pyrin Domain Containing 3 (NLRP3) for example seems to promote inflammasome activation upon diverse DAMP signals, such as excess ATP, NLR Family CARD Domain Containing 4 (NLRC4) senses bacterial type 3 secretion system components and intracellular flagellin (Lamkanfi & Dixit, 2014) (Figure 1). Cytosolic activation and oligomerization of inflammasome associated NLRs promotes Apoptosis-Associated Speck-Like Protein Containing A CARD (ASC) dependent caspase-1 activation, resulting in proteolytic maturation of the cytokines IL1β and IL18 (Rathinam & Fitzgerald, 2016). Caspase-1 furthermore promotes pyroptosis, a rapid form of cell death, exposing cytoplasmic content, including intracellular pathogens, to phagocytes in the periphery (Jorgensen & Miao, 2015). Thus, pyroptosis is fundamentally different from classic, caspase-3/7 induced apoptosis downstream of Cytochrome C leakage from mitochondria or plasma membrane standing death receptors, which culminates in the packaging of cellular content into apoptotic vesicles (Shalini et al., 2015). Cleavage of further protein substrates by caspase-1 impacts on innate defense by affecting cellular metabolism, phagosome maturation and autophagic clearance of intracellular pathogens (Rathinam & Fitzgerald, 2016). The latter is typically activated by autophagy adapters such as the ubiquitin reader Sequestosome-1 or the PRR NOD2, which trigger MAP1A/MAP1B Light Chain 3 (LC3) protein dependent intracellular membrane-engulfment of bacteria and autophagolysosome dependent neutralization (Deretic et al., 2013) (Figure 2). Autophagy activation can in turn dampen inflammasome formation and type I IFN immunity (Deretic et al., 2013), illustrating the complexity of the decision making networks in cell-autonomous antibacterial innate immunity.

Major bacterial recognition pathways in the immune system. TLRs, such as TLR4 can activate NFκB and IRF3 dependent inflammatory mediator and type I IFN production through the MyD88 and TRIF pathway, respectively. NLRs can activate NFκB (e.g., NOD2) or inflammasome-dependent cytokine maturation (e.g., NLRC4). RLRs, such as RigI promote type I IFN production through the MAVS pathway. Type I IFN stimulates STAT-dependent interferon induced gene (ISG) expression via the IFNAR1/2 receptor

lncRNAs in IFNγ and autophagy dependent antibacterial defense. Lymphocytes, such as T or NK cells produce IFNγ in response to phagocyte signals, such as IL12. IFNγ production is promoted by lymphocyte lncRNAs such as IFNG-AS1 or lnc-CD244. Stimulation of infected cells with IFNγ promotes JAK–STAT dependent antibacterial defense via autophagy and nitric oxide production. iNOS production is positively regulated by linc-Cox2. STAT activation is inhibited by Sros1, which is downregulated upon phagocyte activation. LncRNAs MEG3, PCED1B-AS and lincRNA-EPS, which inhibit autophagy, are also downregulated upon bacterial infection
The cellular innate immune response to bacterial pathogens is further shaped by cytokines produced by cells of the adaptive immune system, such as T- and NK-cells. The release of IL12 from PRR-activated macrophages for instance stimulates Th1-polarized CD4+ cells and CD8+ T-cells for type II interferon IFNγ production, which in turn stimulates defense against intracellular bacteria in macrophages and other cell types (Figure 2). IFNγ binding to the IFNγ receptor (IFNGR) activates a JAK/STAT signaling cascade, similar to the above described type I IFN signaling cascade (Schroder et al., 2004). IFNγ induced antibacterial adaptations, for example, include the production of reactive oxygen species or increased presentation of digested bacterial antigens to cytotoxic T cells (Schroder et al., 2004). Furthermore, Th2-polarized CD4+ lymphocytes upon TCR-based antigen sensing produce IL4 and IL13, which stimulate macrophage polarization towards an anti-inflammatory and repair-associated cell type (Atri et al., 2018). Thus, antibacterial immunity relies on an intricate interplay of innate and adaptive immune reactions, promoting bacterial clearance and re-establishment of homeostasis. Imbalances in the innate and adaptive immune pathways can result in attenuated immunity or overshooting inflammatory reactions to bacterial pathogens and organ failure (Biswas & Lopez-Collazo, 2009; D'Elia et al., 2013). Therefore, numerous control mechanisms exist, which promote or negatively regulate the immune response (Biswas & Lopez-Collazo, 2009; D'Elia et al., 2013). Besides protein regulators, noncoding RNAs, including microRNAs (miRNAs) and long noncoding RNAs (lncRNAs), are gaining increasing attention in this context. MiR-146 and miR-155 for instance constitute negative feed-back regulators, limiting TLR4-based immune-activation of mammalian macrophages (Janga et al., 2018; Schulte et al., 2013). Other miRNAs regulate autophagic bacterial clearance or are highjacked by bacterial pathogens to their own advantage (Aznaourova & Schulte, 2021). While the roles of miRNAs in bacterial and viral infections have been studied in detail (K. Pawar, Sharbati, et al., 2016; Schulte et al., 2011; Sen Zhang et al., 2018; Zur Bruegge et al., 2016), research on lncRNAs in antibacterial host-defense is still at its infancy.
Long noncoding RNAs are defined as transcripts with ≥200 nucleotides (nt) that lack protein-coding potential. Advances in high-throughput sequencing technologies in the 2000s have led to the discovery of thousands of lncRNAs in human cells and rodent models, only a fraction of which has been functionally characterized to date (M. Sun & Kraus, 2015). The approximately 20,000 lncRNAs encoded in the human genome constitute a heterogeneous group of enhancer-associated RNAs (eRNAs), intergenic transcripts, sense or antisense transcripts, acting in cis and in trans (Derrien et al., 2012; Kopp & Mendell, 2018). Mechanistically, lncRNAs can act as protein decoys, guides or scaffolds, for example, controlling chromatin factor positioning and thus transcription and genome compaction (Rinn & Chang, 2012). LncRNA PACER for instance decoys the p50 subunit of NFκB and thereby prevents formation of transcriptionally repressive NFκB p50 dimers (Krawczyk & Emerson, 2014). HOTAIR guides the Polycomb Repressive Complex 2 (PRC2) to target promoters to regulate HOX-gene dependent developmental programs (Rinn et al., 2007; Zhao et al., 2008). By bringing PRC2 and the Lysine Demethylase 1A (LSD1)/REST Corepressor (CoREST) complex into special proximity, HOTAIR also serves as a scaffold RNA (Tsai et al., 2010). Besides their important roles in the nucleus, lncRNAs are also increasingly reported to participate in cytoplasmic processes, including translation, protein-ubiquitination or RNA turnover (Aillaud & Schulte, 2020). Thus, unlike miRNAs, lncRNAs do not act through a common biochemical pathway but are mechanistically heterogeneous. Several lncRNAs have recently been described as regulators of sterile inflammatory responses to bacterial PAMPs and cellular defense against live bacterial pathogens.
2 LncRNAS IN STERILE ANTI-BACTERIAL IMMUNITYBoth professional immune cells, such as phagocytes, and tissue-forming cells, such as endothelial and epithelial cells, can initiate NFκB-dependent inflammatory responses to bacterial immune agonists. The defense programs mounted by these cell types differ depending on whether sterile bacterial PAMPs or live bacteria are sensed (see Section 3). Sterile antibacterial immunity can, for example, occur in response to remnants of a cleared infection or injected adjuvants (Hajam et al., 2017). In basic research, sterile bacterial PAMPs or synthetic PRR agonists are used to dissect the participation of immune-regulators in specific immune-signaling pathways. Several recent studies suggest nuclear and cytoplasmic lncRNAs to participate in the positive and negative control mechanisms balancing NFκB-dependent inflammatory responses to bacterial PRR-agonists.
2.1 Positive control of NFκB driven immunity upon PRR-activationPhagocytes, such as monocytes or macrophages, play an essential orchestrating role in the antibacterial innate immune response (Kaufmann & Dorhoi, 2016). In human phagocytes, the cytokine IL1β is among the most highly induced pro-inflammatory mediators upon MyD88-pathway activation by bacterial LPS or lipoproteins. Stimulation of human monocytes with Escherichia coli LPS also induces expression of the nuclear-retained lncRNAs IL-1β-eRNA and IL-1β-RBT46. Both lncRNAs are transcribed from the same gene locus as IL1β and positively regulate IL1β expression (Ilott et al., 2014). In human macrophages, LPS furthermore induces the expression of nuclear lncRNA PACER, which prevents the formation of repressive p50 homodimers and thus promotes NFκB p50/p65 heterodimer formation and immune-gene transcription (Krawczyk & Emerson, 2014). Similar to PACER, lncRNA CARLR upon human macrophage activation with E. coli LPS promotes the expression of immune genes, via direct association with NFκB (Castellanos-Rubio et al., 2017). Importantly, CARLR is conserved and its immune-regulatory function extends to murine phagocytes as well (Castellanos-Rubio et al., 2017). At the level of promoter histone modification, the lncRNAs ROCKi and IL7-AS promote MyD88-dependent gene expression upon human macrophage stimulation with LPS and bacterial lipoprotein mimic Pam3CSK4, respectively (X. Liu et al., 2019; Q. Zhang et al., 2019). Of note, IL7-AS belongs to the few conserved immune-regulatory lncRNAs found both in human and murine cells (Roux et al., 2017).
In murine phagocytes, several lncRNAs with little indication for sequence conservation in human cells (Roux et al., 2017), were found to contribute to PAMP-triggered immunity. In murine macrophages, lincRNA-Tnfaip3 is upregulated in response to LPS and promotes NFκB-dependent gene expression by establishing an activatory High Mobility Group Box 1 (HMGB1)/NFκB complex at immune gene loci (S. Ma et al., 2017). Using an NFκB-GFP reporter system, additional lncRNAs were found to regulate NFκB-dependent immune gene expression in murine macrophages stimulated with bacterial LPS or bacterial lipoprotein analogue Pam3CSK4 (Covarrubias et al., 2017). While partially cytoplasmic lncRNA lincRNA-Cox2 was found to promote degradation of the NFκB inhibitor IκBα in the cytosol, lncRNA AK170409 seemed to be promote NFκB-dependent immune responses in the nuclear compartment (Covarrubias et al., 2017). Besides the positive control of IκBα degradation in the cytoplasm, linc-Cox2 seems to adopt both positive and negative regulatory roles in nuclear immune-gene transcription through association with Heterogeneous Nuclear Ribonucleoprotein hnRNPA/B and A2/B1 complexes (Carpenter et al., 2013) and the SWItch/Sucrose Non-Fermentable (SWI/SNF) chromatin remodeling complex (Hu et al., 2016). Another lncRNA controlling immune gene expression in response to LPS in murine macrophages is AS-IL1α, which acts to recruit RNA polymerase II to the promoter of the neighboring IL1α gene (Chan et al., 2015a). Further lncRNAs, such as FIRRE (Lewandowski et al., 2019; Y. Lu et al., 2017) or lncRNA-CCL2 (Jia et al., 2018) are induced in response to bacterial LPS and seem to promote pro-inflammatory murine phagocyte activation; their precise roles in this context, however, demand further investigation.
Besides pro-inflammatory, PRR-inducible lncRNAs, several immunosuppressive lncRNAs were found to be downregulated upon bacterial PAMP-stimulation, thereby contributing to the progression of the immune response. GAPLINC, for instance, is a functionally and positionally conserved lncRNA which is downregulated upon human and murine macrophage stimulation with various PRR ligands, including bacterial LPS (Vollmers et al., 2021). GAPLINC deficiency enhanced baseline expression of NFκB dependent genes in human cells and mice. This pre-activation of immune-response genes protected GAPLINC-deficient mice from LPS-induced endotoxin shock—a fulminant inflammatory response syndrome, associated with high mortality (Vollmers et al., 2021). Other examples of lncRNAs suppressing premature immune responses, which are downregulated upon bacterial PAMP-stimulation, are lincRNA-EPS (Atianand et al., 2016) and lnc13 in murine phagocytes. While lincRNA-EPS interacts with hnRNPL, lnc13 associates with hnRNPD and Histone Deacetylase 1 (HDAC1). In both cases, lncRNA-protein complex formation counteracts immune gene activation by promoting repressive chromatin states at immune gene promoters (Castellanos-Rubio et al., 2016). In summary, several lncRNAs act in the nucleus and cytoplasm to foster MyD88-NFκB dependent immune gene expression upon activation of human and murine phagocytes by bacterial PRR ligands.
Of note, NFκB-dependent pro-inflammatory immune responses do not only rely on professional immune cells. Other cell types, such as vascular endothelial cells or mucosal epithelial cells vitally contribute to first-line inflammatory responses to bacterial pathogens (Lentsch & Ward, 2000; Luissint et al., 2016). Therefore, it is not surprising that several lncRNAs involved in antibacterial responses of immune cells are also engaged in epithelial immunity. Examples are PACER (Krawczyk & Emerson, 2014), CARLR (Castellanos-Rubio et al., 2017), IL7-AS (X. Liu et al., 2019; Roux et al., 2017), and FIRRE (Y. Lu et al., 2017), which positively regulate both phagocyte and epithelial NFκB-driven sterile immune responses to bacterial PAMPs. Other lncRNAs promote antiviral type I IFN defense pathways in epithelial and endothelial cells (Walther & Schulte, 2020), as well as immune-responses to live bacterial pathogens (see Section 3). Thus, a dense network of lncRNAs grants the induction of leukocyte, epithelial and endothelial host defense in response to bacterial PRR agonists. These cellular reactions need to be restricted to prevent from excessive immune-mediator production and inflammation-associated tissue damage and organ failure (Biswas & Lopez-Collazo, 2009; D'Elia et al., 2013). Several lncRNAs have been implicated in this process, as described in the following section.
2.2 Negative regulation of NFκB driven immunity upon PRR activationIn murine macrophages, exposure to bacterial LPS triggers the induction of several lncRNAs, acting to restrain NFκB-dependent inflammatory responses. Mirt2, for instance, is induced upon TLR4-activation and negatively regulates the TLR-MyD88 pathway by impeding auto-ubiquitination and oligomerization of the signal transduction component TRAF6 (Du et al., 2017). In the nucleus, LPS-induced lncRNA Anti-IL1β inhibits IL1β expression by decreasing histone 3, lysine 4 (H3K4) tri-methylation at the promoter (J. Lu et al., 2013). LncRNA SeT is induced in response to LPS in murine macrophages to suppress the biallelic expression of tumor necrosis factor (TNFα) (Stratigi et al., 2015). In SeT deficient mice, increased TNFα production coincides with increased mortality upon LPS-injection-induced sterile endotoxemia (Stathopoulou et al., 2017). Besides limiting the acute induction of NFκB-dependent immune responses, lncRNAs also assist in the resolution of the inflammatory response to bacterial PAMPs, as evidenced by lncRNA lncFAO. This lncRNA is induced during the late LPS-response of murine macrophages and contributes to inflammation resolution by promoting mitochondrial hydroxyacyl-CoA dehydrogenase beta (HADHB)-dependent fatty acid oxidation, which down-modulates macrophage inflammatory activity (Nakayama et al., 2020).
In a human macrophage model, lncRNA THRIL was reported to be downregulated upon stimulation with bacterial lipoprotein mimic Pam3CSK4. In complex with hnRNPL, THRIL was suggested to promote expression of pro-inflammatory mediator TNFα at the promoter level (Z. Li et al., 2014). Downregulation of THRIL upon macrophage-activation would thus inhibit TNFα production. In an independent RNA-seq study, however, expression of this lncRNA in human phagocytes could not be confirmed (Roux et al., 2017).
Like in phagocytes, other cell types such as epithelial and endothelial cells or fibroblasts employ lncRNAs to dampen NFκB-dependent immune responses. Sensing of bacterial LPS by human intestinal epithelial cells, for instance, leads to the induction of lncRNA TMC3-AS, which negatively regulates NFκB p65 binding to target gene promoters (Ye et al., 2020). In human endothelial cells, lnc-IL7R is induced upon sensing of LPS. Lnc-IL7R limits the inflammatory response by supporting the tri-methylation of H3K27 at immune gene promoters (Cui et al., 2014). In murine fibroblasts, Lethe is induced upon stimulation with TNFα, an inflammation mediator that is, for example, produced by phagocytes upon stimulation with bacterial or viral PAMPs. Lethe interacts with NFκB p65 and inhibits its binding to DNA, thereby restraining NFκB-dependent inflammatory responses (Rapicavoli et al., 2013).
Taken together, lncRNAs vitally participate in the positive and negative cellular control mechanisms preventing from attenuated and overshooting NFκB-dependent immune responses to bacterial PAMPs. Of note, in addition to NFκB-dependent immunity, lncRNAs also participate in the control of type I interferon immunity. The known roles of lncRNAs in this context, however, largely relate to viral infections (Walther & Schulte, 2020). An exception is the nuclear lncRNA LUCAT1, which was recently shown to be rapidly induced following LPS-stimulation of human monocytes, dendritic cells and macrophages, in an NFκB and JAK/STAT dependent manner (Agarwal & Vierbuchen, 2020). Loss- and gain-of-function experiments revealed LUCAT1 to restrain type I interferon stimulated gene expression. This seems to involve LUCAT1 recruitment to chromatin and an inhibitory interaction with STAT1 (Agarwal & Vierbuchen, 2020). The contribution of LUCAT1 to the clearance of life bacterial infections, however, remains to be determined. The already known functions of lncRNAs in the defense against living bacterial pathogens through interferon-activation and other pathways restraining bacterial colonization are described in the following sections.
3 LncRNAS IN HOST INTERACTION WITH LIVE BACTERIAThe adequate activation of antibacterial defense programs by the immune system relies on the discrimination of live bacteria from dead bacteria and sterile PRR agonists. In this context, bacterial RNA was found to act as a viability associated PAMP (vitaPAMP), which triggers a specific TLR8-dependent cytokine response to living bacteria in human antigen-presenting cells (APCs) (Sander et al., 2011; Ugolini et al., 2018). Besides cytokines promoting follicular helper T cell (TFH cell) differentiation and adaptive immunity, this specific APC response to live bacteria also entails prominent induction of the lncRNA linc00346, which so far, however, remains of unknown function (Ugolini et al., 2018). Among the cytokines produced by APCs in response to vitaPAMPs is IL12, which skews naïve CD4+ T-cell differentiation towards a TH1 phenotype and promotes IFNγ production by TH1 cells, CD8+ T cells and NK cells (Kaiko et al., 2008; Ugolini et al., 2018). IFNγ in turn triggers JAK/STAT pathway dependent defense-reactions of APCs and other cell types to live bacteria. This includes increased antigen presentation, production of reactive oxygen species and enhanced autophagic clearance of intracellular bacteria (Ivashkiv, 2018). The adequate activation of IFNγ production by lymphocytes and IFNγ-mediated antimicrobial defense is controlled by several lncRNAs (Figure 2).
3.1 LncRNAs in antimicrobial IFNγ circuitriesIn TH1 cells, CD8+ T-cells and NK cells, IFNγ production is promoted by the lncRNA IFNG-AS1, which is also known as NeST or as Tmevpg1 (Collier et al., 2012; Padua et al., 2016; Petermann et al., 2019; Stein et al., 2019; Vigneau et al., 2003). In murine cells, IFNG-AS1 was shown to promote activating histone methylation events at the IFNγ locus through a direct interaction with the methyltransferase WD Repeat Domain 5 (WDR5) (Gomez et al., 2013). Consequently, mice lacking IFNG-AS1 are more likely to succumb to Salmonella enterica Typhimurium infections than wild-type animals due to reduced IFNγ expression and increased bacterial burden (Gomez et al., 2013). In CD8+ T-cells it was shown that the expression of IFNγ and of the inflammation mediator TNFα is regulated by the T-cell—inhibitory molecule CD244 upon Mycobacterium tuberculosis infection (Y. Wang et al., 2015). CD244 induces the expression of a lncRNA, lncRNA-CD244, which directly interacts with the PRC2 subunit Enhancer of Zeste Homolog 2 (EZH2) to suppress IFNγ and TNFα expression at the chromatin-level (Y. Wang et al., 2015).
In phagocytes, stimulation with IFNγ activates the IFNGR-JAK–STAT signaling pathway, which was found to be regulated by lncRNA Sros1 in the context of L. monocytogenes infection (Xu et al., 2019). Mechanistically, L. monocytogenes upregulates miR-1, which targets cytoplasmic Sros1 for degradation. This in turn was suggested to release Sros1-dependent suppression of the RNA binding protein Cell Cycle Associated Protein 1 (CAPRIN1), which stabilizes Stat1 mRNA and increases STAT1 protein levels and thus IFNγ signaling. Thereby, inducible degradation of Sros1 contributes to clearance of intracellular bacteria (Xu et al., 2019). IFNγ also contributes to the polarization of macrophages towards a pro-inflammatory M1-like phenotype, as opposed to a rather anti-inflammatory phenotype induced by IL4. Knockdown of linc-Cox2 in murine RAW264.7 macrophages was shown to suppress the clearance of intracellular Mycobacterium bovis BCG (Bacillus Calmette–Guérin) by inhibiting M1 polarization and nitric oxide production (W. Chen et al., 2019). Vice versa, linc-Cox2 overexpression enhanced binding of NFκB to the inducible Nitric Oxide Synthase (iNOS) promoter, thereby propelling nitric oxide production (W. Chen et al., 2019). Beyond linc-Cox2, in Mycobacterium tuberculosis infected murine macrophages, IFNγ pre-stimulation was found to pronouncedly alter the lncRNA expression profile (Roy et al., 2018). The relevance of this observation in the context of the course of bacterial infection, however, remains to be determined.
3.2 LncRNAs in autophagic clearance of bacteriaAutophagy is a central process in the maintenance of cellular homeostasis, promoting lysosomal breakdown of proteins or damaged organelles (Islam Khan et al., 2018). Autophagy also critically contributes to defense against intracellular bacteria, such as M. tuberculosis or the vaccine strain M. bovis BCG (Gutierrez et al., 2004) and is enhanced upon IFNγ stimulation (Ivashkiv, 2018). Recently, lncRNAs were found to play critical roles in control of autophagy under diverse pathological conditions (Bao et al., 2017; Z. Liu et al., 2016; Yang, Zhang, Li, & Liu, 2016c), including bacterial infections (Figure 2).
Maternally expressed lncRNA gene 3 (MEG3), for example, was shown to be significantly downregulated in IFNγ-treated THP1-derived macrophages challenged with M. bovis BCG. Knockdown of MEG3 fostered the induction of autophagy and promoted clearance of M. bovis (Pawar, Hanisch, Palma Vera, Einspanier, & Sharbati, 2016a). Interestingly, different from M. bovis infection, MEG3 is upregulated in human THP1-derived macrophages upon challenge with M. smegmatis, which is considered nonpathogenic, but not with M. avum, which is a facultative pathogen (Sharbati et al., 2019). Thus, MEG3 might play different roles in response to different mycobacterial species. PCED1B-AS is another lncRNA involved in the regulation of autophagy during mycobacterial infection. In CD14+ monocytes from patients with active tuberculosis PCED1B-AS levels were reduced, accompanied by decreased apoptosis and enhanced autophagy (M. Li et al., 2019). Mechanistically, PCED1B-AS was suggested to bind miRNA-155 and impair its function (M. Li et al., 2019). The downregulation of miR-155 targets Forkhead Box O3 (FOXO3) and Ras Homolog, MTORC1 Binding (RHEB) due to PCED1B-AS1 downregulation was suggested to increase LC3 expression and reduce caspase-3 levels, potentially explaining the observed effects on autophagy and apoptosis, respectively (M. Li et al., 2019). In response to microbial PAMPs, macrophages downregulate the long intergenic RNA-erythroid pro-survival (lincRNA-EPS), which functions as a repressor of immune gene expression (Atianand et al., 2016). Similar to PCED1B-AS1, lincRNA-EPS expression was found to be decreased in monocytes from patients with active pulmonary tuberculosis (PTB) (Ke et al., 2020). Knockdown of lincRNA-EPS inhibited apoptosis and enhanced autophagy in M. bovis BCG-infected murine RAW264.7 macrophages by promoting JNK/MAPK signaling (Ke et al., 2020). Taken together, the clearance of intracellular bacteria, such as M. tuberculosis, M. bovis or L. monocytogenes through the process of autophagy is tightly controlled by lncRNAs, directly impacting on autophagic activity of infected host cells or on the responsiveness to and production of autophagy inducer IFNγ.
3.3 LncRNAs in inflammatory responses to living bacteriaAs already noted above, the type II interferon IFNγ not only promotes autophagy but also potentiates cellular immune responses to PRR ligands. The resulting production of inflammation mediators, such as IL1β, Cox2, IL6, or TNFα critically contributes to bacterial clearance but also to inflammatory pathologies if not properly controlled (Van Amersfoort et al., 2003). To avoid from attenuated or overshooting inflammation during antimicrobial immune responses, intricate regulatory RNA mechanisms have evolved, controlling the PRR-NFκB signaling pathways (Walther & Schulte, 2020). The nuclear lncRNA AS-IL1α, for instance, is upregulated upon L. monocytogenes infection in an NFκB dependent manner (Chan et al., 2015b). AS-IL1α in turn promotes the expression of IL1α by recruiting RNA polymerase II to the IL1α promotor (Chan et al., 2015b). Another lncRNA upregulated in murine macrophages upon L. monocytogenes infection is linc-Cox2 (Carpenter et al., 2013), which positively and negatively regulates distinct sets of immune genes through nuclear and cytoplasmic mechanisms (see Section 2.1). In human peripheral blood mononuclear cells (PBMCs) from patients with tuberculosis and in THP1-derived macrophages infected with M. tuberculosis Nuclear Paraspeckle Assembly Transcript 1 (NEAT1) was found to be upregulated compared to control group cells (Huang et al., 2018). Knockdown of NEAT1 in THP1-derived macrophages led to reduced IL6 but not TNFα levels after infection with M. tuberculosis and increased bacterial load (Huang et al., 2018). While these observations remain to be explained mechanistically, other studies using sterile PRR ligands found NEAT1 to displace the paraspeckle protein and transcriptional regulator SFPQ (Splicing Factor Proline And Glutamine Rich) from target immune gene promoters (Imamura et al., 2014; H. Ma, Ming, et al., 2017). An lncRNA known to be downregulated upon PRR-stimulation in phagocytes is lincRNA-EPS, which relieves hnRNPL-dependent suppression of immune gene expression at the promoter level (see Section 2.1). In line with these observations, in L. monocytogenes challenged macrophages and dendritic cells from lincRNA-EPS−/− mice increased levels of pro-inflammatory cytokines, such as TNFα and IL6 were detected (Agliano et al., 2019). In addition, increased expression of iNOS and increased nitric oxide production was observed. Consequently, bacterial colony forming units (CFUs), recovered from liver and spleen of L. monocytogenes infected lincRNA-EPS−/− mice, were reduced compared to wild-type animals (Agliano et al., 2019). Thus, PRR-dependent downregulation of lincRNA-EPS upon phagocyte infection serves to promote pro-inflammatory immune responses and bacterial clearance. In human blood-derived macrophages infected with the gram-negative bacterial pathogen Legionella pneumophila, upregulation of the lncRNAs Macrophage Interferon-regulatory lncRNA 1 (MaIL1) and Linc01215 was observed (Aznaourova et al., 2020). MaIL1 is induced in a TLR4-NFκB dependent manner, but regulates TLR4-TRIF dependent type I interferon production. Thus, different from the lncRNAs depicted above, MaIL1 bridges the TLR-NFκB and the TLR-TRIF pathway. Mechanistically, MaIL1 associates with the ubiquitin reader and signaling adapter OPTN to promote OPTN and TBK1-kinase dependent phosphorylation of the IRF3 transcription factor and thus type I IFN expression. MaIL1 silencing in macrophages increased intracellular L. pneumophila CFU counts, which could be reversed by the addition of recombinant type I IFN (Aznaourova et al., 2020). Taken together, several lncRNAs regulate the production of inflammation mediators and type I interferons in bacterially infected human and murine phagocytes in an NFκB dependent manner (Figure 3), thereby contributing to productive antibacterial defense.

LncRNAs in inflammatory pathway activation during bacterial infection. Upon innate detection of bacteria, such as L. monocytogenes, M. tuberculosis, P. aeruginosa or L. pneumophila, several lncRNAs are regulated, which impact on pro-inflammatory and type I IFN gene expression. Through PRRs, lncRNAs AS-IL1α and linc-Cox2 are up-, whereas lincRNA-EPS is downregulated to promote NFκB pathway activation. Of note, linc-Cox2 can also suppress the activation of a set of immune genes. LncRNA MEG3-4 is a positive regulator of pro-inflammatory cytokine IL1β expression and is downregulated upon detection of bacteria. NEAT1v2, which promotes immune gene expression, is upregulated in response to intracellular, replicating S. Typhimurium. MaIL1 is an lncRNA, which is upregulated through the MyD88-NFκB pathway but interacts with components of the TRIF-pathway to promote type I IFN expression and IFN-dependent antimicrobial defense
In addition to phagocyte immunity, lncRNAs also regulate inflammatory reactions of epithelial cells and heterogeneous mucosal cell networks to living bacterial pathogens. Pseudomonas aeruginosa, for instance is an opportunistic gram-negative pathogen, which can cause severe lung infections, especially in immune-compromised individuals (Sadikot, Blackwell, Christman, & Prince, 2005). RNA-seq analysis of P. aeruoniosa infected bronchial epithelial cells from cystic fibrosis (CF) patients compared to non-CF epithelial cells revealed 108 lncRNAs that were differentially expressed over the course of the infection. Two lncRNAs, MEG9 and BLACAT1, were validated to be significantly downregulated in CF cells after infection (Balloy et al., 2017). In lung tissue of P. aeruginosa infected mice, another member of the maternally expressed gene family, the lncRNA MEG3, was found to be significantly downregulated in a TLR4-NFκB signaling dependent manner (R. Li & Fang, 2018). Downregulation of MEG3 transcript isofom 4 reduced IL1β expression, whereas MEG3-4 overexpression was found to promote excessive inflammation and propel bacterial colonization of murine lungs in vivo. Mechanistically, lncRNA MEG3-4 was suggested to suppress IL1β by competing with IL1β mRNA for miR-138 binding (R. Li & Fang, 2018). Many additional lncRNAs were shown to be regulated at the epithelial and endothelial barriers. Using a 3D reconstructed model of the human intestinal barrier, dozens of uncharacterized lncRNAs were shown to be regulated upon Salmonella enterica Typhimurium infection in the communicating intestinal epithelial cells, endothelial cells, monocytes and NK cells (Schulte, 2020). Similarly, in human brain microvascular endothelial cells (hBMECs) dozens of lncRNAs were up- or downregulated following infection with meningitic E. coli (R. Yang et al., 2016a). The roles of these lncRNAs in bacterial infection and host defense, however, remain to be determined. Taken together, phagocyte, epithelial and endothelial lncRNAs seem to vitally participate in the control of pro-inflammatory immune responses to live bacterial pathogens, with relevance to inflammation- and infection-induced pathologies.
3.4 LncRNAs linking bacterial infection to cancerIn the context of colorectal cancer (CRC), Fusobacterium nucleatum was identified as a gut microbe particularly enriched in the local tumor tissue environment (C. H. Sun et al., 2019). RNA-seq analysis of CRC cells exposed to F. nucleatum revealed 43 lncRNAs, which were upregulated in response to the bacterium, in addition to classic immune response genes such as CCL20 or IL8. Among these lncRNAs was Keratin 7 antisense RNA (KRT7-AS), which was induced in an NFκB dependent manner (S. Chen et al., 2020). Loss and gain of function assays revealed KRT7-AS to promote metastasis formation by CRC cells in nude mice by upregulating KR
Comments (0)