
Remember me
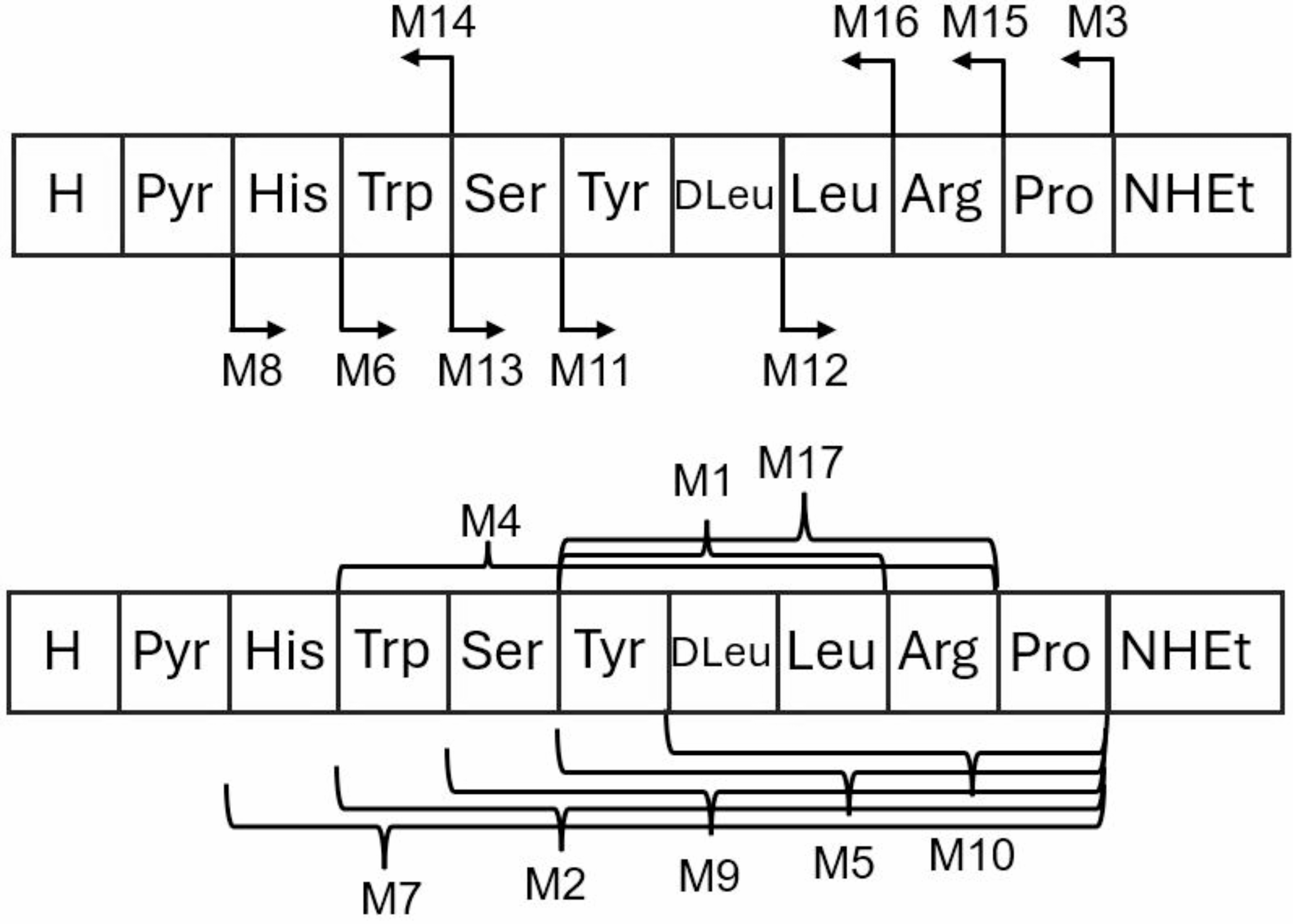


The Uy234 and QnCs-Buap peptides were assayed for their antimicrobial activities against S. enterica serovar Typhimurium SL1344, A. baumannii ATCC 17,978, K. pneumoniae ATCC 700,603, S. aureus ATCC 29,213, ATCC 43,300 (MRSA), 6N3, 18F1, B. subtilis 168, E. gallinarum 55-1228, E. mundtii ATCC E-5, and E. faecalis 24,212. We tested 100 µM for each peptide and specifically 503.67 µM (Uy234) and 711.38 µM (QnCs-Buap), respectively. Under these conditions, no significant inhibition was found with the disc diffusion assay but with the well diffusion one, a significant inhibitory effect on the growth of S. aureus ATCC 29,213 was achieved with the same concentrations of each peptide. The inhibition halos obtained were quantified at 11 mm (QnCs-Buap) and 12 mm (Uy234) (Fig. 1).
Fig. 1
Antimicrobial activity of Uy234 and QnCs-Buap peptides using the well diffusion assay. (A) S. enterica serovar Typhimurium SL1344, (B) A. baumannii ATCC 17,978, (C) K. pneumoniae ATCC 700,603, and (D) S. aureus ATCC 29,213. Well 1: QnCs-Buap (711.38 µM): Well 2: QnCs-Buap (100 µM); Well 3: Uy234 (503.67 µM); Well 4: Uy234 (100 µM); Well 5: sterile water; Well 6: Water-Acetonitrile (1:4); Well 7: 15% CinnAcar (cinnamon extract)
As a growth inhibitory effect was observed for the S. aureus ATCC 29,213 strain, which is susceptible to antibiotics, we wondered whether both peptides could inhibit antibiotic-resistant S. aureus strains. Hence, the activity of these peptides was tested against methicillin-resistant S. aureus ATCC 43,300 (MRSA), as well as against S. aureus 6N3 and 18F1 strains, classified as multidrug-resistant (MDR). Notably, the Uy234 peptide induced inhibition halos on all three S. aureus MDR strains using the lowest dose (100 µM); in contrast, no inhibition halos were observed with the QnCs-Buap peptide (Fig. 2). These results indicate that the Uy234 peptide is biologically active, especially against MDR, MRSA strains of S. aureus but less active against A. baumannii.
Fig. 2
Antimicrobial activity of the Uy234 and QnCs-Buap peptides against S. aureus strains by the well diffusion assay. (A) S. aureus ATCC 43300 (MRSA); (B) S. aureus 6N3 (MDR); (C) S. aureus 18F1 (MDR). Well 1: Uy234 (503.67 µM); Well 2: Uy234 (100 µM); Well 3: QnCs-Buap (711.38 µM); Well 4: QnCs-Buap (100 µM); Well 5: Sterile water; Well 6: 1:4 water-acetonitrile; Well 7: 15% CinnAcar (cinnamon extract)
The MIC and the MBC for both peptides, Uy234 and QnCs-Buap, were determined against the eleven strains studied (Table 2). As expected, the QnCs-Buap peptide, whose net charge is + 1 but corresponds to the charge of the N-terminal end, showed little antimicrobial effect against all the strains studied. On the contrary, the Uy234 peptide, with a net charge of + 3 (+ 2 in terms of side-chain charges) was particularly effective against MDR and MRSA strains of S. aureus but less efficient against (A) baumannii and (B) subtilis. The effective dose in these cases was 50 and 25 µM, respectively. This result agrees quite well with what was observed in the well-diffusion assay (see Supplementary Fig. S1). Remarkably, the Uy234 scorpion peptide exhibited bactericidal activity at low doses (12.5 µM) against the four S. aureus strains evaluated, and it also showed bactericidal activity against B. subtilis 168, at 50 µM, although this same treatment was only able to inhibit the growth of A. baumannii (Table 2). Taken together, these data demonstrate that only the Uy234 peptide possesses antimicrobial activity, particularly against S. aureus strains of medical interest.
Table 2 Minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of the Uy234 and QnCs-Buap peptidesHemolytic Activity in Human ErythrocytesThe hemolytic activities of both peptides were characterized using fresh red blood cells. Since this characterization was previously done in a previous work (Cesa-Luna et al. 2019), we now decided to calculate the hemolytic effect using mixtures of both peptides. As depicted in Fig. 3, the combination of both peptides showed a low hemolytic activity, reaching only 2.5 ± 0.7% cell lysis at 50 µM of each peptide, which is four times the concentration of the MBC at which the Uy234 peptide exhibits bactericidal activity against all the S. aureus strains tested. In any case, the hemolytic activity in these conditions was not higher than 3.5%, which is consistent with the HC25 value reported for this peptide, i.e. HC25 = 370 µM, which is the concentration needed to induce 25% hemolysis (Cesa-Luna et al. 2019). It can therefore be useful to compare the MIC in sensitive bacteria and the HC25 to estimate therapeutic indices of 7.4 (A. baumannii), 14.8 (B. subtilis), and 59.2 (S. aureus). These findings reinforce the notion that the Uy234 peptide is highly effective against multidrug-resistant strains of S. aureus and is relatively harmless against human cells.
Fig. 3
Hemolytic activity of the Uy234/QnCs-Buap peptide combination. Hemolytic activity against human red blood cells after 1 h of exposure to the Uy234/QnCs-Buap combination is shown in red squares. The green circles show the negative hemolysis control, in which 1X PBS buffer was applied. As a positive cell lysis control, 10% Triton X-100 was tested, causing 100% cell lysis
Structural Analysis and Conformational DynamicsTo investigate in more detail the prominent antimicrobial activity that we described for the peptide Uy234 against S. aureus, we evaluated in detail the physicochemical and structural parameters of this peptide, focusing on the contribution of the amide group and net charge. With the calculation of these physicochemical parameters (Table 3) it is possible to have an estimate of the lytic potential of each peptide (Table 4).
Fig. 4
Helical wheel projections and structures of Uy234 (A) and QnCs-Buap (B) peptides. Left panels show the carboxilated form. N and C red letters indicate the N- and C-ends, respectively. Amino acids in blue color are positively charged, and residues in yellow are hydrophobic. Polar serines are depicted in violet and kink-promoting prolines in green. The black arrows indicate the hydrophobic moment µH. Right models depict the 3D hydrophobic moments of C-end amidated peptides. Black arrows show the 3D hydrophobic moment vector and colors are equivalent
In Fig. 4 we depict helical wheel projections and structures for the two modeled peptides, as well as the magnitude of their µH and 3DHM hydrophobic moments. As can be noted, although both peptides have an equivalent hydrophobic ratio (Table 3), the Uy234 peptide, in its carboxylated (Fig. 4A) or amidated form (Fig. 4B), has a significantly lower hydrophobic vector compared to the one present in the QnCs-Buap peptide. Indeed, the θ angle of this vector is most optimally oriented in the QnCs-Buap consensus peptide, which is consistent both with their Wimley-White and the Boman indices, significantly better for this peptide (Table 4). However, the absence of positive charges in side chains and the lower electrostatic energy of the QnCs-Buap peptide allow us to affirm that this peptide is not capable of overcoming the energy barrier during the first phase of interaction with the membrane surface, i.e. the interfacial adsorption stage (Kabelka and Vácha 2021), and probably it preferably remains in the aqueous milieu. Uy234, on the contrary, by possessing two contiguous positive charges in flexible side chains and possessing better electrostatic energy, is capable of overcoming this energetic barrier and transitioning to an ordered state, which facilitates the translocation of the peptide through the lipid bilayer.
Table 3 Physicochemical parameters of Uy234 and QnCs-Buap peptidesTable 4 Wimley-White and Boman indices for Uy234 and QnCs-Buap peptidesThen, to evaluate the contribution of the presence of the terminal amide group in both peptides, we first estimated the main flexibility of both peptides in their carboxylated forms. As can be noted, the main flexibility of the Uy234 peptide (mBf = 1.793) is significantly higher than that present for the QnCs-Buap consensus peptide (mBf = 1.657). Furthermore, it is interesting to note that both the dipole moment (DM), determined by the geometry of the amino and carbonyl functional groups of each peptide unit in the main chain and aligned in parallel with the axial Z-axis of the α-helix (Hol et al. 1978), as well as the internal bending of the peptide along the same Z-axis are significantly different between the amidated Uy234 peptide (164 ± 8.14 D, κf = 19o) and QnCs-Buap peptide (172.6 ± 2.19 D, κf\(\:\approx\:\) 1°) (Table 5). Overall, these data suggest greater conformational freedom for peptide Uy234.
Table 5 Flexibility determinants for Uy234 and QnCs-Buap amidated peptidesHaving noted this significant difference in terms of the intrinsic flexibility in both peptides, we set out to evaluate in atomic detail the effect of the presence of this amide group in each peptide. To do this, we compared and contrast both structures by evaluating both the H-bonding pattern and the torsion effect promoted by the central proline in both carboxylated and amidated peptides. In Supplementary Fig. 2 we show the three regions of interest for the structural analysis of these peptides: (1) the hydrophobic N-terminal end, rich in Phe and with the presence of a second proline in position 2 for the peptide Uy234; (2) the central proline whose presence would facilitate a bending effect in both peptides; and (3) the last four residues of the C-terminal end, where the amide group is located, a sequence with the presence of two basic residues [-IKRL-], charged at neutral pH in the Uy234 peptide, and highly flexible, whereas in the consensus QnCs-Buap peptide the corresponding region [-SGLI-] is significantly more rigid (Fig. 5).
Fig. 5
Intrinsic flexibility of Uy234 and QnCs-Buap peptides. Comparison between the intrinsic flexibility of Uy234 (blue, mBf = 1.793) and QnCs-Buap (red, mBf = 1.675) peptides in terms of the normalized B-factor (nBf)
Once the three regions of each peptide were established to study in atomic detail, we set out to evaluate the impact that the presence of the terminal amide group has on the structure of both peptides. To do this, we compared the H-bonding pattern inside the main chain of each model and calculated the internal bending angle (κf) along the axial axis of each peptide. These results are depicted in Fig. 6 and Supplementary Fig. S3 where it is shown that the presence of the amide group promotes the formation of a denser H-bonding network inside the main chain of each peptide, notably Uy234, which is intrinsically more flexible.
Fig. 6
Role of C-terminal amidation in Uy234 and QnCs-Buap peptides. (A) Role of amidation in the Uy234 peptide by promoting the formation of a network of double H-bonds and reducing the internal bending of the peptide (κf) from 32º to 19º, which is facilitated by the presence of Pro 9. (B) The amidation of the QnCs-Buap peptide has a smaller effect in terms of the H-bonding network (yellow dashes) and internal bending given its greater intrinsic rigidity. The presence of the amide group also reduces specific interatomic distances (blue and purple dash lines)
The high intrinsic flexibility of the Uy234 peptide in its carboxylated form is significantly reduced when the amide group is present. This creates new H-bonds in the main chain, reconfiguring the helical structure and promoting two factors that affect its flexibility: (i) DM increases and, in consequence, (ii) reduction in the internal bending angle (κf), around the central proline of each peptide, is facilitated. Regarding QnCs-Buap peptide, given its more hydrophobic character and lower intrinsic flexibility (Fig. 5), the effect of amidation is less pronounced in terms of these two structural parameters (Table 5).
Finally, intending to explore the effect of the greater flexibility and net charge of the Uy234 peptide, factors that seem to be particularly determining for the lytic activity of this peptide compared to QnCs-Buap, we carried out a series of MD simulations of each peptide in aqueous solution. These results are shown in Fig. 7 and represent 100 ns using the GROMACS forcefield on the WebGRO server. We evaluate two structural parameters: (1) the root mean square deviation (RMSD) of the backbone concerning an ordered bent structure for each peptide in its amidated form, and (2) the root mean square fluctuation (RMSF), which estimates the trajectory of each residue in the chain and whose value depends largely on the type and intrinsic flexibility of each lateral group. On the other hand, we also evaluate three electrostatic parameters that depend predominantly on the interaction of each peptide with the aqueous solvent: (1) the average number of H-bonds over time, (2) the radius of gyration (Rg) which provides information for the conformational freedom of each peptide under the evaluated conditions, and (3) the solvent-accessible surface area of each peptide.
Fig. 7
RMSD, RMSF, H-bonding evolution, Rg and SAS of Uy234 and QnCs-Buap peptides. Comparison of the root mean square deviations (RMSD) of the Cα backbone atoms, root mean square fluctuations (RMSF), H-bonding evolution, radius of gyration (Rg), and solvent-accessible surface (SAS) area of Uy234 (blue) and QnCs-Buap (red) peptides at T = 300 K
As expected, being significantly more hydrophilic and flexible, the Uy234 peptide interacts better in the aqueous phase, which is manifested by a higher RMSD than the inactive QnCs-Buap peptide. In addition, Uy234 generates more H-bonds on average and the SAS area is also slightly higher (~ 20 nm2) compared to the consensus peptide (~ 14 nm2). The Rg for Uy234 (0.74 ± 0.06 nm) and QnCs-Buap (0.66 ± 0.05 nm) are also distinct, in good agreement with more side-chain mobility in terms of RMSF for Uy234, in comparison with the inactive peptide (Fig. 7). Despite its increased mobility, the Uy234 peptide remains rather ordered in the N-terminal region, just upstream of the central proline (Pro9); this feature could be determinant for its bioactivity.
Having noticed this, and to further study the possible effect on the degree of ordering in both peptides, we decided to evaluate the RMSF parameter in detail by increasing the temperature. In the Supplementary Fig. S4 we show that by increasing the temperature to 305 K, the degree of disorder of the Uy234 peptide is equivalent to the one of QnCs-Buap peptide at room temperature (300 K). Indeed, we observed that in control conditions (300 K) some β-structures are formed, which is consistent with an Rg similar to that of the short and inactive QnCs-Buap peptide. Notably, at 300 K the QnCs-Buap peptide is virtually entirely disordered. This slightly greater degree of order at 300 K for the Uy234 peptide (Movie-1) compared to QnCs-Buap (Movie-2) is shown in the two attached movies as supplementary material. Finally, the number of interatomic contacts was also estimated, and they confirmed a high degree of order in the Uy234 peptide at room temperature, in comparison with the innocuous QnCs-Buap peptide in similar conditions. These data reinforce the notion that, despite having similar degrees of compaction, the active peptide Uy234 remains predominantly ordered in the aqueous phase, forming stable secondary structures.
Comments (0)