
Remember me

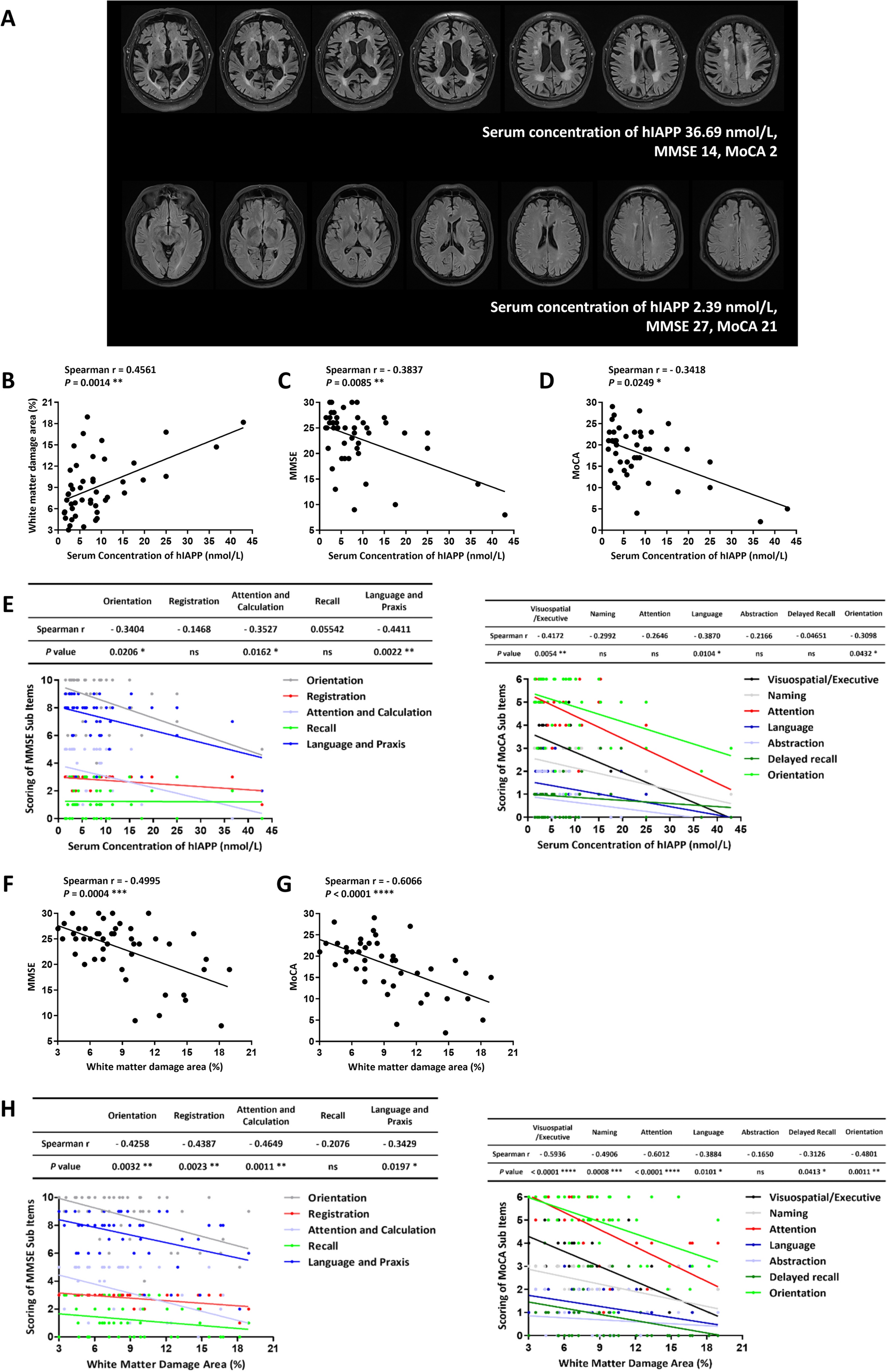

To assess the spontaneous behavior patterns and characteristics of Shank3b mutant mice, we employed a 3D-motion capture system and Behavioral Atlas software [7] (Fig. 1A) to collect and analyze behavioral data from 114 Shank3b mutant mice aged between 12–16 weeks (male: KO = 20, HE = 15, and WT = 19; female: KO = 21, HE = 12, and WT = 17) from both sexes. To accurately capture the motion of different parts of the body, we tracked 16 key body parts to reconstruct the skeleton of a mouse. Using unsupervised clustering, 40 types of behavioral movements were identified. Then, we manually designated 13 types of major behavioral movements based on highly similar postures, including running, trotting, walking, right turning, left turning, stepping, rearing, climbing, hunching, rising, sniffing, grooming, and pausing. Behavioral atlas analysis effectively revealed the structure of the spontaneous behavior in Shank3b mutant mice (Fig. 1B, C) (Table 1).
Different Genotypes Exhibit Different Spontaneous Behavioral Characteristics in Both SexesTo analyze the spontaneous behavioral repertoires of Shank3b WT, HE, and KO mice, we examined the behavioral sequences (Fig. 2A, Fig. S1A, B) and analyzed time fractions (Fig. 2B) and frequencies (Fig. 2C) of all the movements in the three groups for both sexes over 60 min. Our findings revealed that walking and stepping, which may represent motor deficit function in patients, showed significant differences across the three groups for both sexes (Fig. 2C), and KO mice of both sexes exhibited a significant reduction in other movements such as trotting, right turning, climbing, rising, and sniffing, which may represent the exploratory function in patients, but an increase in grooming, which may represent the stereotyped movements in patients, compared to their WT counterparts. Female KO mice also showed significantly reduced running and increased hunching, which may represent forced posture in patients, compared to the WT group. Interestingly, HE mice from both sexes showed a significant decrease in walking, stepping, and sniffing compared to the KO mice, coupled with a pronounced increase in pausing compared to WT mice and in grooming compared to female KO mice. Furthermore, we evaluated sex differences in all movements across the three groups and found that the frequency of the walking exhibited significant differences between sexes within each genotype (Fig. 2D, Fig. S1C). Specifically, the WT groups displayed the most pronounced differences, followed by the HE group, while the KO group revealed no noticeable differences.
Fig. 2
Distinct spontaneous behavioral characteristics of Shank3b mutant mice in both sexes over 60 min. A Representative ethogram of the five significant different movements among Shank3b WT, HE, and KO mice of both male (left) and female (right) mice (Up, WT; Middle, HE; Down, KO). Five animals were randomly selected in each group. Middle color-coded images correspond to the five different movements, top to bottom: walking (orange), stepping (light pink), hunching (blue), grooming (rose red), and pausing (gray). B Comparison of the fraction of thirteen movements among Shank3b WT, HE, and KO mice (Left: male; KO, deep green; HE, pale green; WT, blank; Right: female; KO, dark red; HE, pink; WT, blank). Middle color-coded labels indicate the movements. Statistics: two-way ANOVA followed by Turkey post hoc multiple comparisons test. C Comparison of the frequency of thirteen movements among Shank3b WT, HE, and KO mice (Left: male; Right: female). Statistics: two-way ANOVA followed by Turkey post hoc multiple comparisons test. D Comparison of the frequency of thirteen movements between male and female Shank3b mutant mice (left: WT; Middle: HE; Right: KO). Statistics: two-way ANOVA followed by Turkey post hoc multiple comparisons test. ***P < 0.001, **P < 0.01, *P < 0.05.
Besides, the total travel distance in the open field arena showed that HE and KO mice of both sexes exhibited a significant decrease in walking compared to the WT group (Fig. S2), with the KO group decreasing most prominent. No significant differences were observed between male and female mice of the same genotype group (Fig. S2E–G). Overall, our findings indicated that different genotypes of Shank3b mutant mice exhibited diverse movement characteristics across both sexes.
Genotypes Show Distinct Temporal Patterns of Spontaneous BehaviorsTo better understand the functional behavior characteristics of Shank3b WT, HE, and KO mice, we classified 13 movements into 5 clusters based on their similarities or specific links to ASD, including locomotion, exploration, maintenance, forced posture, and nap. Similarly, we analyzed the time fractions and frequencies of each cluster across both sexes over 60 min. Our results showed that time-frequency among all three groups for both sexes showed significant differences in locomotion, with the KO group of the most prominent, mainly attributed to walking and stepping (Fig. 3A, B, I). Further, KO mice exhibited a significant decrease in exploration of both sexes and prominent increases in maintenance and nap for the female group, probably resulting from the increased climbing, grooming, pausing, and decreased sniffing (Fig. 3A, B, I). Additionally, sex differences in the movement fractions (Fig. S3) and frequencies across time (Fig. 3C) of each cluster were examined across the three genotypes. We found that locomotion and exploration of both sexes exhibited significant differences among the three groups. Subsequently, we divided the observational 60-min period into six 10-min segments and conducted an in-depth analysis of the temporal patterns of each cluster (Fig. 3D-H) and movement (Fig. 3I) for both sexes (Fig. S3, S4). Our results showed that both male and female KO mice allocated less time for locomotion cluster across all 10-minute segments, particularly during the last three segments for male HE mice compared with the WT (Fig. 3D). Female KO mice spent significantly more time in the maintenance cluster from the third segment onwards but were only limited to the second segment for the male KO group (Fig. 3E). The notable increase in time spent on the nap cluster of female KO and male HE groups in the whole 60-min period was largely attributed to the significant increase during the last two segments for females and the fourth and final segments for males, respectively (Fig. 3H). No noticeable differences in exploration and forced posture were found in each segment except between the second to fourth segments for the female KO group (Fig. 3F, G). Besides, we conducted a time-based comparison of sex differences in the time spent on the 5 clusters (Fig. S5) and 13 movements between male and female Shank3b WT (Fig. S6), HE (Fig. S7), and KO (Fig. S8) mice. We found that throughout the entire duration, as well as across all six 10-minute segments, there were virtually no significant sex differences observed, indicating a mild sex-dependent variation in the temporal behavioral characteristics among the three groups. These findings provide further insights into different temporal behavioral characteristics among the three groups of Shank3b mutant mice of both sexes.
Fig. 3
Temporal dynamics of clusters exhibited by both male and female Shank3b mutant mice. A–B Comparison of the time fractions and frequency of five clusters among male (A left: fractions; Right: frequency) and female (B left: fractions; Right: frequency) Shank3b WT, HE, and KO mice over 60 min. Statistics: two-way ANOVA followed by Turkey post hoc multiple comparisons test. All data are presented as means ± SEM. C Comparison of the frequency of thirteen movements between male and female Shank3b mutant mice (left: WT; Middle: HE; Right: KO). Statistics: two-way ANOVA followed by Turkey post hoc multiple comparisons test. ***P < 0.001, **P < 0.01, *P < 0.05. D–H Temporal dynamics of the five clusters in male and female Shank3b mutant mice over 60 min, with the sequence from D to H representing locomotion, maintenance, forced posture, exploration, and nap. The upper part of the panels shows the results for male mice (KO, deep green; HE, pale green; WT, blank), while the lower part shows the results for female mice (KO, dark red; HE, pink; WT, blank). Statistics: two-way ANOVA followed by Dunnett post hoc multiple comparisons test. All data are presented as means ± SEM. I Temporal dynamics of the 13 movements in every 10-min interval of male (left) and female (right) Shank3b mutant mice. Statistics: two-way ANOVA followed by Dunnett post hoc multiple comparisons test. ***P < 0.001, **P < 0.01, *P < 0.05.
Additionally, we explored the temporal characteristics of the distance traveled by the six groups. Although there was a significant increase in the total distance traveled across all groups and both sexes across all segments, we found a significant decline at each segment for the KO groups and during the last two segments for the HE groups compared to the WT groups (Fig. S9A–D). Notably, the distance traveled in each segment was significantly decreased over time among the three groups, with the most drastic decline being in the KO groups across all segments, while only the fourth and fifth for the male HE groups and the third to fifth segments for female HE groups compared with the WT group, respectively (Fig. S9E–H).
Spatial Preferences in the Open Field Vary Among Different GenotypesThe spatial preferences in the open field test are commonly used to evaluate the anxiety-like behavior in rodents. Typically, the center area is defined as half the length of the open field, and decreased time spent in the center is considered as an increase in anxiety-like level, as shown in Shank3b KO male mice [34, 35]. With the new division method, we found that the length of the square boundaries (‘a’, 136.24 mm, 136.98 mm, and 134.94 mm for WT, HE, and KO mice, respectively) did not differ significantly among WT, HE, and KO mice (Fig. 4A, B). Therefore, we standardized the length at 135 mm for subsequent analysis. The time spent in different areas and types of movements therein were evaluated for Shank3b mutant mice using both traditional and data-driven region division methods. Comparative analyses of two divisions showed that the KO mice of both sexes spent less time in the central area but more time in the perimeter area compared to the WT group (Fig. 4C–H). The same analysis revealed that male KO mice had decreased walking in the centre and decreased pausing in the perimeter area, while female KO mice had decreased walking, stepping, and right turning in the centre and increased hunching, grooming, and pausing in the perimeter area based on the new division. However, only grooming movement significantly increased in the perimeter area in male KO mice, but no noticeable movement significantly decreased in the center area by the traditional division. These findings revealed that our data-driven method is more compatible with the animal's actual trajectory than the traditional ones, suggesting our region division method provides a more detailed insight into behaviors.
Fig. 4
Spatial preference characteristics of movements exhibited by male and female Shank3b mutant mice. A Density distribution curve with back coordinates of three Shank3b mutant mice, the red triangle denoting the starting point of the 90% peak prominence as a division of the heterogeneity occupancy density regions. B Representative images of the spatial preference of three Shank3b mutant mice, with the dotted lines delineating the boundary between the center and the perimeter. C, F Comparison of the time fractions in the center and the perimeter among male (C) and female (F) Shank3b WT, HE, and KO mice in 60 min. D, G Comparison of the time fractions per unit area in the center and the perimeter among male (D) and female (G) Shank3b WT, HE, and KO mice in 60 min. E, H The pie diagrams compare the time percentage of the center and perimeter area for male (E) and female (H) Shank3b WT, HE, and KO mice. I Comparison of movement variation in the center (left) and perimeter (right) areas for male Shank3b WT, HE, and KO Mice Over a 60-min duration. J Comparison of movement variations in the center (left) and perimeter (right) areas for female Shank3b WT, HE, and KO mice over a 60-min duration. ***P < 0.001, **P < 0.01, *P < 0.05.
Different Genotypes Display Distinct Behavioral Transition PatternsNext, we investigated the influence of the genotypes and sexes on the behavioral temporal sequence of Shank3b mice, we analyzed the transition patterns within clusters over an hour. The results showed that the most significant changes and differences among the three groups in both sexes in transition patterns were primarily observed during the first half (Fig. 5A, B). We observed a gradual increase in mutual transition between maintenance and nap across the groups from WT to HE to KO, of both sexes, while the opposite trends were observed between maintenance and locomotion, as well as between forced posture and locomotion (Fig. 5C, D). Moreover, to elucidate the significance of clusters in a behavioral transition network, we computed two-time segments. Our data showed that, during the initial time segment, clusters for both sexes functioned as transitional hubs, manifesting distinct transition patterns when compared to those observed in the WT group. In addition in the HE group, forced posture and nap in males, along with maintenance in females similarly displayed divergent transition patterns relative to WT mice. For example, both the male and female KO mice could transition from locomotion to exploration, as well as to hunching, while locomotion will only transform to exploration in WT mice (Fig. 5E, F). Consistent with the transition patterns of each cluster in the initial time segment, the transition pattern for each cluster as a transitional hub was less pronounced than that in the latter 30-min period (Fig. 5G–J). Overall, mice exhibited relatively stable behavioral connections and transitional patterns, yet transition strategies and behavioral organization of KO and HE groups varied from the WT group.
Fig. 5
Behavioral transition probabilities of five clusters in male and female Shank3b mutant mice. A, B Behavioral similarities between the first and the second 30-min duration for male (A) and female (B) Shank3b mutant mice (Left: WT; Middle, HE; Right: KO), with each cell represented the overall similarity of each group within the corresponding timeframes. C, D, G, H Behavioral transition probabilities during the first and the second 30 min across the five clusters in male (first: C; second: G) and female (first: D; second: H) Shank3b WT (left), HE (middle), and KO (right) mice. The five color-coded circles denote the five corresponding clusters. The size of each circle indicates the relative occurrence probability of each cluster, with larger circles denoting higher probabilities. The connecting lines among every two circles represent the transition probabilities of the two clusters, with greater line thickness indicating a higher transition rate. Lines around the circles indicate the self-transition probability based on the movements grouped into the clusters. E, F, I, J Connection properties of each cluster in betweenness centrality for both male (first: E; second: I) and female (first: F; second: J) Shank3b WT, HE, and KO mice during the second 30 min. The magnitude of transition probabilities is symbolized by the length and/or thickness of the lines linking two nodes, with greater length and/or thickness denoting higher probabilities. All lines representing transition probabilities of less than 0.05 are intentionally omitted.
Characterization Data Models Distinguish among Different GenotypesWe explored the feasibility of distinguishing autism-associated mice from WT mice based on the altered spontaneous behavioral patterns exhibited by Shank3b WT, HE, and KO mice. LDA was employed to integrate and analyze the behavioral data of all 13 movements. As expected, Shank3b mutant mice were largely separated from distinct genotypes for both sexes (Fig. 6A, E). We then trained several other LDA classifiers on different parameters that were traditionally applied to discriminate autism, such as grooming, traveling distance, position in the arena, and clusters, to predict genotypes of different groups (Fig. 6B, D, F, H). Our results revealed that prediction accuracy of the classifier based on movement fractions and frequency (discrimination: male, 1.0; female, 0.92) outperformed the classification based on grooming (discrimination: male, 0.48; female, 0.72), distance (discrimination: male, 0.59; female, 0.56), position in the open field arena (discrimination: male, 0.57; female, 0.57), and clusters (discrimination: male, 0.68; female, 0.68) in identifying autistic from WT animals (Fig. 6C, G). This highlights that the classifier trained on 13 movements exhibited more critical insights compared with other behavioral parameters used in most previous methods.
Fig. 6
Behavioral dissimilarity among Shank3b WT, HE, and WT across both sexes. A, E Low-dimensional representation of the thirteen movements in male (A) (WT, blank; HE, pale green; KO, deep green) and female (E) (WT, blank; HE, pink; KO, dark red) Shank3b mutant mice using LDA linear model. B, F Normalized classification matrices by all thirteen movements (across rows and columns) for distinguishing male (B) and female (F) Shank3b WT, HE, and KO mice. The numerical values within the matrices represent the count of true and/or predicted instances for each group. The color bar is shared by all matrices. The values, ranging from 0 (depicted in white) to 1 (depicted in black) indicate increasing discrimination. An ideal classifier performance corresponds to a diagonal black with otherwise white fields (Discrimination of 1). C, G The discrimination of different classifiers for male (C) and female (G) Shank3b WT, HE, and KO mice. D, H Normalized classification matrices by traditional classifiers for distinguishing male (D) and female (H) Shank3b WT, HE, and KO mice.
Comments (0)