
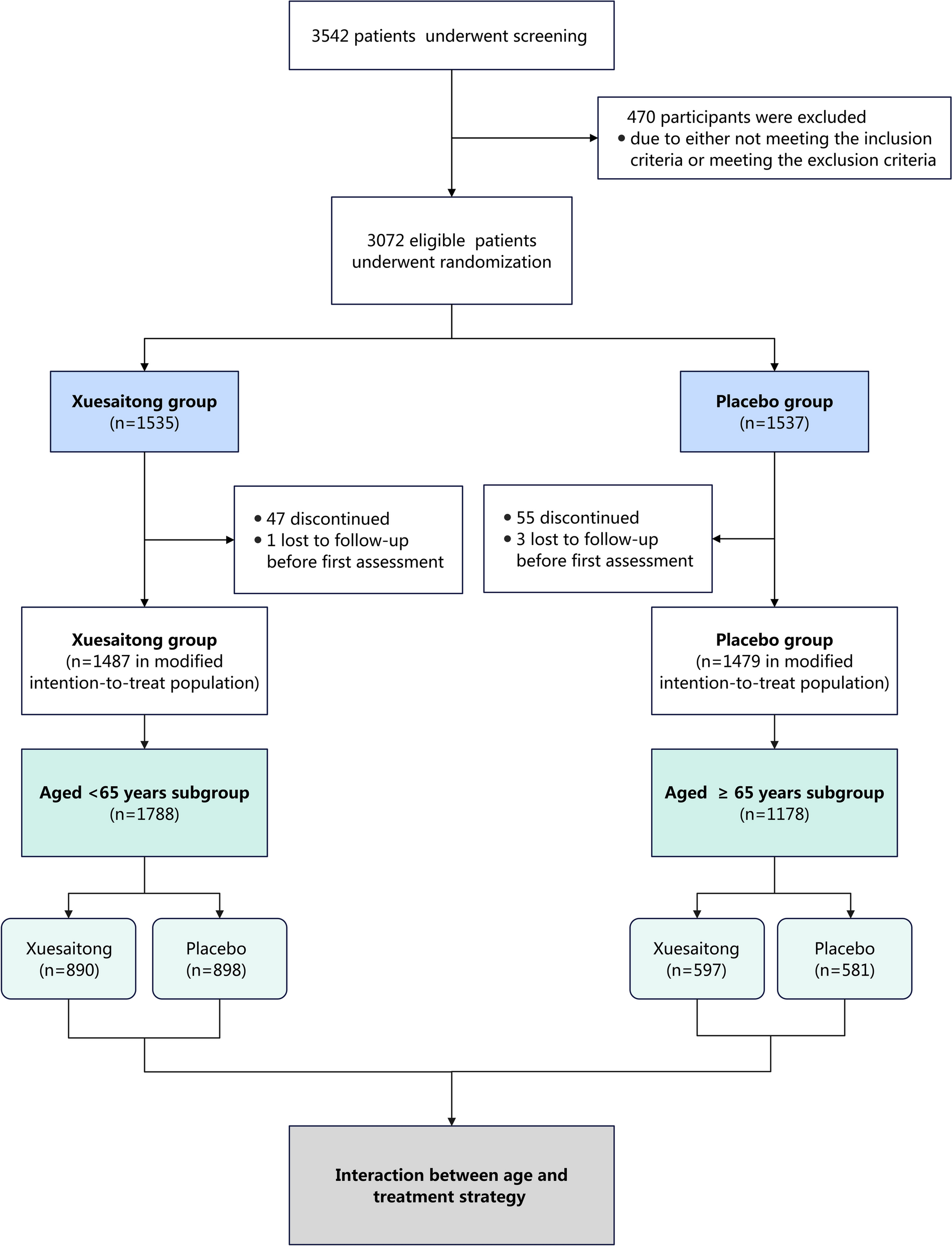
Remember me








DOX-induced cardiotoxicity is a classical chronic cardiotoxic model [2, 3]. The mice received a low dose of DOX (5 mg/kg/week) intraperitoneally for 5 weeks. Based on the therapeutic outcomes, a dose of Lig (20 mg/kg/day) and TMZ (10 mg/kg/day) were selected for the treatment of DOX-induced cardiotoxicity in mice for 1 month. TMZ is used as a clinical treatment for cardiac diseases by improving myocardial energy homeostasis and cardiac systolic-diastolic function; therefore, we served as a positive control drug [23]. After treatment, efforts were made to assess the anti-atrophy effect of Lig. The results revealed that the heart weight-to-body weight ratio decreased in DOX-treated mice compared to saline-treated mice, while it increased in Lig-treated mice (Fig. 1A and B). Furthermore, the cardiotoxicity of DOX was evaluated using biochemical markers associated with heart failure. The data showed a significant increase in the concentrations of CK-MB and LDH after DOX treatment. However, following treatment with Lig and TMZ, the concentrations of CK-MB and LDH decreased, with noteworthy emphasis on Lig's superior therapeutic effect compared to TMZ (Fig. 1C and D). Cardiac function was assessed by echocardiography, focusing on ejection fraction (EF) and shortening fraction (FS). Compared with mice treated with normal saline, EF and FS in DOX-treated mice were significantly reduced, indicating that DOX-induced cardiac dysfunction in mice. Treatment with Lig and TMZ resulted in significantly higher EF and FS compared to the DOX group (Fig. 1E–G). Additionally, HE staining and Masson trichrome staining revealed improvements in myocardial inflammatory infiltration, expansion of intercellular spaces, and fibrosis in DOX-treated mice after treatment with Lig and TMZ (Fig. 1H). These findings suggest that Lig can ameliorate cardiac dysfunction in DOX-induced mice and improve structural changes associated with heart failure. In our previous study, we have also confirmed that Lig has positive muscle strength in rats with heart failure [19]. Subsequently, we conducted tests to verify this phenomenon in heart failure rats, where the left anterior descending coronary artery was ligated, followed by 8 weeks of Lig gavage (Fig. s1A). In hemodynamic assessments, we observed a gradual improvement in left ventricular systolic pressure (LVSP), left ventricular end-diastolic pressure (LVEDP), +dp/dt, and −dp/dt with increasing Lig dosage (Fig. s1B–E). Further tests revealed a gradual decrease in the levels of nitric oxide (NO) and free fatty acids (FFA) under Lig treatment (Fig. s1F and G). These preliminary results suggest that Lig plays a certain role in the treatment of heart failure.
Fig. 1
Lig improves DOX-induced cardiac dysfunction in mice. A, B The gross heart pictures and the HW/BW ratio of mice (n = 10). C, D Effects of Lig on serum levels of CK-MB and LDH in mice caused by DOX (n = 6). E–G The LVEF and LVFS were measured by echocardiography in each group (n = 6). H HE and Masson staining images of heart tissues in mice (n = 3). I, J MDA and SOD levels in mice caused by DOX (n = 6). K Effects of Lig on heart levels of ROS caused by DOX (n = 3). Data are presented as the mean ± SD. **p < 0.01, ***p < 0.001 compared with control group; #p < 0.05, ##p < 0.01 compared with model group
Considering that mitochondrial oxidative stress plays a crucial role in the development of cardiac dysfunction under various pathological conditions [24]. The levels of ROS, MDA, and SOD in the myocardial tissue of mice were assessed. ROS tissue fluorescence analysis indicated an increase in ROS expression after DOX treatment. However, the ROS content in the heart decreased after treatment with Lig and TMZ. Regarding MDA and SOD, MDA content increased, and SOD activity decreased in the hearts of the DOX group, while MDA content decreased, and SOD activity increased under the treatment of Lig (Fig. 1I and J). Additionally, oxidative damage in rats was evaluated based on MDA, SOD, and ATP contents, and the results were consistent with the DOX-induced chronic cardiotoxic model in mice (Fig. s1H–J). ROS has the potential to induce apoptosis by affecting mitochondrial metabolism [25]. Western blotting analysis revealed that Lig treatment decreased cardiomyocyte apoptosis by reducing the Bax protein level and increasing the Bcl-2 protein level (Fig. s2A). Furthermore, immunofluorescence analysis demonstrated that apoptosis-inducing factor (AIF) was more expressed in the DOX group, but after treatment with Lig and TMZ, AIF expression decreased, with Lig exhibiting a better therapeutic effect than TMZ (Fig. s2B). These results collectively suggest that Lig can counteract oxidative stress damage and apoptosis induced by cardiotoxicity in vivo.
Lig activates AMPK/SIRT3 signaling and alleviates pyroptosis mediated by Caspase-3/GSDME in DOX-induced cardiac toxicityIn recent years, numerous studies have highlighted that DOX-induced myocardial injury involves multiple biological processes, including oxidative stress, lipid peroxidation, DNA damage, mitochondrial injury, apoptosis, and autophagy. Among these processes, AMPK has emerged as a pivotal regulator of endogenous defense mechanisms against various pathological processes in the heart [26, 27]. To confirm whether AMPK and SIRT3 signaling are involved in Lig’s protection against DOX-induced myocardial injury in vivo, Fig. 2A demonstrates that Lig treatment significantly increased p-AMPK levels and promoted SIRT3 protein expression. As AMPK is a key regulator of biological energy metabolism, consistent with the aforementioned results, cardiac ATP content decreased in the DOX group and significantly increased after Lig treatment compared with the DOX group (Fig. 2B). In heart failure rats, we also observed significant increases in p-AMPK and SIRT3 under Lig treatment (Fig. S1K). Hence, Lig may represent an effective prevention and treatment method by activating the AMPK/SIRT3 signaling pathway against DOX-induced cardiotoxicity.
Fig. 2
Lig improves DOX-induced cardiac toxicity in mice by activating AMPK/SIRT1 signaling and alleviating pyroptosis mediated by caspase-3/GSDME. A Representative western blotting images and statistical results of p-AMPK, AMPK and SIRT3 in mice (n = 3). B Effects of Lig on heart levels of ATP in mice caused by DOX (n = 5). C, D The serum levels of IL-1β and IL-18 of mice measured by ELISA. E The expression of GSDME-N, cleaved-caspase-3, caspase-3, IL-1β and IL-6 (n = 3). Data are presented as the mean ± SD. **p < 0.01, ***p < 0.001, ****p < 0.0001 compared with control group; #p < 0.05, ##p < 0.01, ###p < 0.001 compared with model group
Activation of AMPK/SIRT3 signaling has been reported to induce pyroptosis [28]. Pyroptosis is recognized as a programmed cell death regulated by inflammation. Serum levels of inflammatory factors IL-1β and IL-18 increased in the DOX group, whereas they decreased significantly after treatment with Lig (Fig. 2C and D). GSDME, specifically cleaved by caspase-3, releases its N-terminal pore-forming domain under certain apoptotic stimuli, leading to pyroptotic death. In the DOX group, GSDME and Cleaved-caspase-3 protein levels increased; however, it was reversed after Lig treatment (Fig. 2E). Similarly, in heart failure rats, GSDME and Cleaved-caspase-3 protein levels were significantly reduced after treatment with Lig (Fig. S1K). The expression of related inflammatory factors IL-1β and IL-6 also decreased after Lig treatment compared with those in the DOX group (Fig. 2E). NF-κB has pro-inflammatory effects and is associated with pyroptosis. Western blot results showed that NF-κB activation increased in the DOX group and decreased NF-κB activation after Lig intervention (Fig. S2A). Collectively, these results indicate that Lig significantly improves AMPK/SIRT3 signaling and reduces pyroptosis in vivo.
Lig improves DOX-induced H9C2 cell viability and decreases mitochondrial oxidative stress by activating AMPK/SIRT3 signalingIndeed, DOX can induce both acute and chronic cardiotoxicity, manifesting as various cardiovascular complications. These complications include tachycardia, arrhythmia, hypotension, transient depression of left ventricular function, and in severe cases, refractory late-onset cardiomyopathy [29]. To investigate whether Lig has a protective effect against DOX-induced cardiotoxicity in vitro, we initially examined its impact on cell viability. As depicted in Fig. 3A, compared with the control group, the viability of H9C2 cells treated with various concentrations of Lig did not exhibit significant changes within 24 h. This indicates that Lig did not exert significant toxicity on H9C2 cells at the experimental concentrations. In comparison to the DOX (3 μM) treatment group, different doses of Lig were found to enhance the viability of H9C2 cells, with Lig (100 μM) significantly improving cell viability and morphological damage in H9C2 cells (Fig. 3B and C). Subsequently, intracellular ROS production was analyzed based on ROS-mediated conversion of non-fluorescent 2′,7′-DCFH-DA to fluorescent DCFH, which exhibited enhanced fluorescence following the production of reactive metabolites [24]. In H9C2 cells treated with DOX, there was a significant increase in ROS production, and Lig treatment demonstrated a more effective inhibition of ROS production compared to TMZ-treated H9C2 cells (Fig. 3D). Similarly, we detected cellular ROS production by flow cytometry with the same results as in Fig. 3D (Fig. S3A). Additionally, MDA content increased, and SOD activity decreased in DOX-treated cells, which were reversed after Lig treatment (Fig. 3E and F). ATP levels exhibited a similar trend in cells (Fig. 3G). Similarly, levels of Bax and Bcl-2 were detected by western blotting, revealing an increase in Bax protein and a decrease in Bcl-2 protein levels in DOX-induced H9C2 cells. After Lig treatment, the Bax protein level decreased, and the Bcl-2 protein level increased (Fig. 3H). Hoechst can penetrate the cell membrane and bind to DNA, and in apoptotic cells, Hoechst-stained nuclei appear in a more pronounced blue light. Hoechst staining further confirmed that apoptosis of H9C2 cells treated with DOX was evident, and this apoptosis significantly decreased after Lig incubation (Fig. 3J and Fig. S3B). Collectively, these data suggest that Lig improves DOX-induced viability and reduces oxidative stress in H9C2 cells.
Fig. 3
Lig decreased DOX-induced H9C2 cells viability inhibition, mitochondrial oxidative stress, and cell apoptosis by improving AMPK/SIRT3 signaling. A Cytotoxicity of Lig on H9C2 cells (n = 5). B Effects of Lig on the cell viability of H9C2 cells induced by DOX (n = 5). C The cellular morphology of H9C2 cells in bright image (n = 6). Scale bar represents 100 µm. D Intracellular ROS level in H9C2 cells (n = 4). Scale bar represents 50 µm. E–G Intracellular MDA, SOD and ATP level in H9C2 cells (n = 3). H, I The expression of Bax, Bcl-2, p-AMPK, AMPK and SIRT3 in H9C2 cells by western blotting assays (n = 3). J Representative images of Hochest staining (n = 6). Scale bar represents 50 µm. K The level of MMP was measured by JC-1 assay kit (n = 6). Scale bar represents 25 µm. Data are presented as the mean ± SD. **p < 0.01, ***p < 0.001 compared with control group; #p < 0.05, ##p < 0.01, ###p < 0.001 compared with model group
Consistent with the findings in vivo, which demonstrated that Lig stimulates AMPK/SIRT3 signaling and mitigates DOX-induced cardiac toxicity damage, we also assessed the protein levels of AMPK and SIRT3 in H9C2 cells. The results revealed that the protein expression levels were consistent with those observed in mice (Fig. 3I). The fundamental role of AMPK is to maintain mitochondrial health, and SIRT3 exerts regulatory effects on the structure and function of mitochondria. Mitochondrial dysfunction can lead to a dissipation of MMP and an increase in ROS. Therefore, we utilized the JC-1 assay and the data indicated that the membrane potential of H9C2 cells treated with Lig and TMZ significantly increased, with the membrane potential of Lig-treated cells higher than that of the TMZ group (Fig. 3K). These results suggest that Lig plays a myocardial protective role against DOX-induced cytotoxicity by activating AMPK and SIRT3 signaling pathways to improve mitochondrial function.
Lig inhibits pyroptosis-associated proteins caspase-3 and GSDME in DOX-treated H9C2 cellsMitochondrial dysfunction may be involved in caspase-3/GSDME-mediated cell pyroptosis. At the morphological level, we observed that H9C2 cells exhibited increasing pyroptotic membrane ballooning and took up PI red after DOX treatment. This effect was reversed with Lig and TMZ treatment, as observed in PI staining and flow experiments (Fig. 4A and D). Furthermore, pyroptosis leads to increased LDH release, and the results showed that Lig significantly decreased DOX-induced LDH release in H9C2 cells (Fig. 4B). Interestingly, these bubble-like protrusions, termed pyroptotic bodies, were previously observed in cells undergoing pyroptosis. As a distinct form of programmed cell death, pyroptosis can be distinguished from apoptosis by morphological features, where the latter shows cell shrinkage, cytoplasmic condensation, and the formation of apoptotic bodies with varying sizes around the nucleus [30]. It is noteworthy that pyroptosis is more prominently observed in cells upon treatment with DOX compared to Lig treatment (Fig. 4C). Subsequently, we tested the levels of pyroptosis-associated proteins, and the results indicated that the expressions of GSDME, Cleaved-caspase-3, IL-6, IL-1β and NF-κB proteins increased in DOX-treated H9C2 cells. However, these increases were significantly reversed after Lig treatment (Fig. 4E and Fig. S3C). These results demonstrate that Lig has the capability to inhibit pyroptosis in DOX-treated H9C2 cells.
Fig. 4
Lig treatment inhibits the caspase-3/GSDME signaling pathway in DOX-treated H9C2 cells. A Representative images of PI staining (n = 6). Scale bar represents 50 µm. B Effects of Lig on intracellular LDH level in H9C2 cells treated by DOX (n = 3). C Representative scanning electron microscopy (SEM) images showed the morphological changes of cultured H9C2 under different treatments (n = 3). D H9C2 cells stained with PI were detected by flow cytometry (n = 3). E The protein expression of GSDME-N, cleaved-caspase-3, caspase-3, IL-6, IL-1β in H9C2 cells (n = 3). Data are presented as the mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 compared with control group; #p < 0.05, ##p < 0.01 compared with model group
Activation of AMPK/SIRT3 signaling is required for Lig to reduce the mitochondrial respiration and pyroptosis in cardiomyocytesTo further elucidate the relationship between Lig and AMPK/SIRT3 signaling, we employed the AMPK inhibitor (CC) and specific AMPK-siRNA to assess cytotoxicity in DOX-induced H9C2 cells. Western blotting analysis confirmed the effective inhibition of siAMPK in H9C2 cells (Fig. S6A). Our findings revealed that CC treatment did not inhibit cell viability, but it significantly prevented the protective effects of Lig in DOX-treated H9C2 cells. This suggests that AMPK/SIRT3 signaling is essential for Lig to mitigate DOX-induced cytotoxicity in H9C2 cells (Fig. S3). Moreover, the cell morphology exhibited similar characteristics in Fig. 5A. As a crucial indicator of mitochondrial damage, DCFH-DA serves as a general ROS probe, highlighting the burst of ROS after stimulating cells with 3 μM DOX. However, this effect was mitigated by Lig treatment and then reversed with the AMPK inhibitor CC or siAMPK treatment (Fig. 5B and Fig S6B). The results of ROS by flow cytometry detection were consistent with the results of ROS fluorescence (Fig. S5A). ROS can induce modifications in related proteins in respiratory chain complexes and even interfere with DNA and RNA replication, affecting their structure and function at the transcriptional level. This cascade of events can lead to oxidative respiratory chain dysfunction and, ultimately, cellular energy metabolism disorders [31]. Subsequently, we utilized JC-1 to assess MMP, and observed that the green (monomeric) fluorescence was increased in the DOX treatment group compared to the Lig treatment group (Fig. 5C). Moreover, treatment with AMPK inhibitor CC or siAMPK reversed the effect of Lig and increased green fluorescence (Fig. 5C and Fig S6C). This result suggests that mitochondrial dysfunction may be a critical factor in the cell death processes induced by DOX, in which Lig reverses H9C2 cell death and AMPK plays an important role.
Fig. 5
Activation of AMPK/SIRT3 signaling is required for Lig to reduce the mitochondrial respiration and pyroptosis in cardiomyocytes. A Morphology in H9C2 cells. Scale bar represents 100 µm (n = 6). B Intracellular ROS level in H9C2 cells (n = 4). Scale bar represents 50 µm. C MMP level was measured by JC-1 assay kit. Scale bar represents 25 µm (n = 6). D Effects of Lig and CC on intracellular ATP level in H9C2 cells (n = 3). E Representative images of Hochest and PI staining (n = 6). Scale bar represents 50 µm. F The expression of SIRT3, Bax and Bcl-2 in H9C2 cells treated by DOX (n = 3). G H9C2 cells stained with PI were detected by flow cytometry (n = 3). H The protein level of GSDME-N, cleaved-caspase-3, caspase-3, IL-6, IL-1β in H9C2 cells. (n = 3) I Intracellular LDH level in H9C2 cells treated by DOX (n = 3). Data are presented as the mean ± SD. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 compared with control group; #p < 0.05, ##p < 0.01 compared with model group. ns indicates no significance
Furthermore, as MMP is crucial for ATP generation, we treated H9C2 cells with Lig and CC to investigate whether Lig would affect ATP levels. The results demonstrated that samples treated with Lig exhibited significantly higher ATP activity compared to the DOX group, indicating that Lig could enhance ATP release. However, this effect was diminished after AMPK inhibition (CC), suggesting that the improvement of mitochondrial function by Lig was attenuated by inhibiting AMPK (Fig. 5D). Mitochondria play a pivotal role in regulating apoptosis signals. Functional damage to mitochondria can result in an increase in MMP, leading to the leakage of internal substances from mitochondria and the activation of apoptosis [32]. Hoechst staining confirmed that the anti-apoptotic effect of Lig was compromised after combining Lig with CC, and PI staining indicated an increase in the DOX treatment group (Fig. 5E, Fig. S5B and S6D). Similarly, western blotting analysis revealed an increase in Bax expression and a decrease in Bcl-2 levels after AMPK inhibition, indicating that inhibiting AMPK weakened the anti-apoptotic effect of Lig (Fig. 5F and Fig. S6E). As an indicator of pyroptosis, increased LDH release was detected in H9C2 cells after AMPK inhibition, and the PI staining results were consistent with the above findings (Fig. 5G and I and Fig. S6D). When combined with CC, the inhibitory effect of Lig was mitigated, as evidenced by increased expression levels of GSDME-N and Cleaved-caspase-3 (Fig. 5H). Similarly, in combination with siAMPK, as with CC, expression levels of GSDME-N and Cleaved-caspase-3 increased significantly (Fig. S6F). Inhibition of AMPK activity using CC cotreatment counteracted Lig's anti-apoptotic effects by elevating the cell apoptotic index. These results underscore the involvement of AMPK activation in Lig's anti-apoptotic effect against DOX-induced cardiotoxicity.
Knockdown of GSDME enhances the protective effect of Lig against DOX-induced H9C2 cell apoptosisTo validate the role of GSDME-mediated pyroptosis in the protective action of Lig, we employed specific GSDME-siRNA to knock down GSDME. Western blotting analysis confirmed the effective suppression of GSDME in H9C2 cells, achieving a transfection efficiency of approximately 25% (Fig. S7A). GSDME can be used as a key protein in apoptosis and pyrogenic transformation of cells. However, when GSDME is overexpressed, it will not only cause pyrogenic death of cells but also cause mitochondria to release pro-apoptotic factors to further cause apoptosis [33]. Following, combined treatment with siGSDME, the apoptosis induced by DOX in H9C2 cells was markedly reduced (Fig. S7B). Subsequently, we delved into the expression of apoptosis-related proteins in H9C2 cells. The western blotting assays unveiled that Lig treatment was linked to an upregulation in Bcl-2 expression and a downregulation in Bax expression compared to the DOX treatment group. Intriguingly, when combined with siGSDME treatment, the apoptosis of H9C2 cells was further diminished (Fig. S7C). The occurrence of pyroptosis leads to the release of a large number of inflammatory factors. Consequently, the expression levels of inflammation-related proteins IL-6 and IL-1β witnessed a further decrease in the Lig combined with siGSDME group compared to the Lig-alone group (Fig. S7C). Collectively, these findings suggest that GSDME knockdown may augment the protective efficacy of Lig against DOX-induced cytotoxicity in H9C2 cells.
Effect of Lig treatment combined with AMPK inhibitors on cardiac toxicity, dysfunction, mitochondrial oxidative stress injury and pyroptosis on DOX-stimulated mice heartsDOX has served as a pivotal component in chemotherapy for over five decades, forming the foundation of treatment protocols for various pediatric and adult cancers. However, its cardiotoxic effects have emerged as a growing and significant clinical challenge confronted by cardiologists [2, 3]. Previously, DOX-related mitochondrial dysfunction has become a likely pathological mechanism for DOX-induced cardiotoxicity, which is also a major reason ROS generation caused by DOX results in irreversible myocardial oxidative injury. In line with prior investigations, when the total cumulative dose of DOX reached 25 mg/kg, echocardiography demonstrated that CC nullified the beneficial impact of Lig on enhancing cardiac function in DOX-exposed mice (Fig. 6A–C). Combining Lig with CC led to a corresponding reduction in heart weight/body weight ratios (Fig. 6D and E). In addition, myocardial damage indexes CK-MB and LDH showed an increase after CC treatment, but there was no significant difference between CK-MB and Lig group. (Fig. 6F and G). This trend was also evident in HE and Masson tricolor staining, where CC annulled the positive effects of Lig, exacerbating inflammatory infiltration and collagen deposition in the myocardium of DOX-exposed mice (Fig. 6H). In summary, these findings underscored that Lig alleviated DOX-induced cardiac dysfunction, and when combined with AMPK inhibitor CC, CC partially reverses the protective effect of Lig, making Lig virtually ineffective in vivo.
Fig. 6
Effects of Lig combined with AMPK inhibitor on cardiac structure and function in DOX-induced mice. A–C The LVEF and LVFS were measured by echocardiography in each group (n = 6). D, E The gross heart pictures and the HW/BW ratio of mice (n = 10). F, G Effects of Lig and CC on serum levels of CK-MB and LDH in mice (n = 6). H HE and Masson staining images of heart tissues (n = 3). Data are presented as the mean ± SD. **p < 0.01, ***p < 0.001 compared with control group; #p < 0.05, ##p < 0.01 compared with model group. ns indicates no significance
Considering that mitochondrial oxidative stress is the primary contributor to DOX-induced cardiomyopathy, we investigated the oxidative stress parameters in myocardial tissue. The co-administration of Lig with CC significantly attenuated the antioxidative stress effect of Lig on the heart subjected to DOX stimulation. Compared to the Lig treatment group, there was an increase in ROS and MDA contents, a decrease in SOD activity, and a significant reduction in ATP content after treatment with Lig combined with CC (Fig. 7A–D). Additionally, the co-treatment with Lig and CC resulted in increased cardiomyocyte apoptosis, counteracting the anti-apoptotic effect of Lig (Fig. 7E). Tissue immunofluorescence of AIF revealed a significant increase in the combination treatment with CC (Fig. 7F). In Fig. s5A, it is evident that CC nullifies the effect of Lig on the reduction of GSDME-N and cleaved-caspase-3. Furthermore, inflammation-related factors were counteracted by CC, as evidenced by increased levels of IL-6 and IL-1β proteins, and the expression of IL-1β and IL-18 in the serum was consistent with these findings (Fig. s5B and C). Collectively, these results suggest that Lig can alleviate DOX-induced cardiotoxicity, reduce myocardial oxidative stress and apoptosis through the AMPK/SIRT3 signaling pathway, and inhibit the GSDME/caspase-3 pathway to mitigate DOX-induced pyroptosis of cardiomyocytes.
Fig. 7
Effect of Lig treatment combined with AMPK inhibitors on cardiac mitochondrial oxidative stress injury and pyroptosis in mice. A The heart levels of ROS in mice caused by DOX (n = 3). B–D The heart levels of MDA, SOD and ATP in mice (n = 6). E The protein expression of SIRT3, Bax and Bcl-2 in mice treated by DOX (n = 3). F Immunofluorescence detects the levels of AIF on cardiac (n = 3). Data are presented as the mean ± SD. **p < 0.01, ***p < 0.001 compared with control group; #p < 0.05, ##p < 0.01, ###p < 0.001 compared with model group. ns indicates no significance
Comments (0)