
Remember me

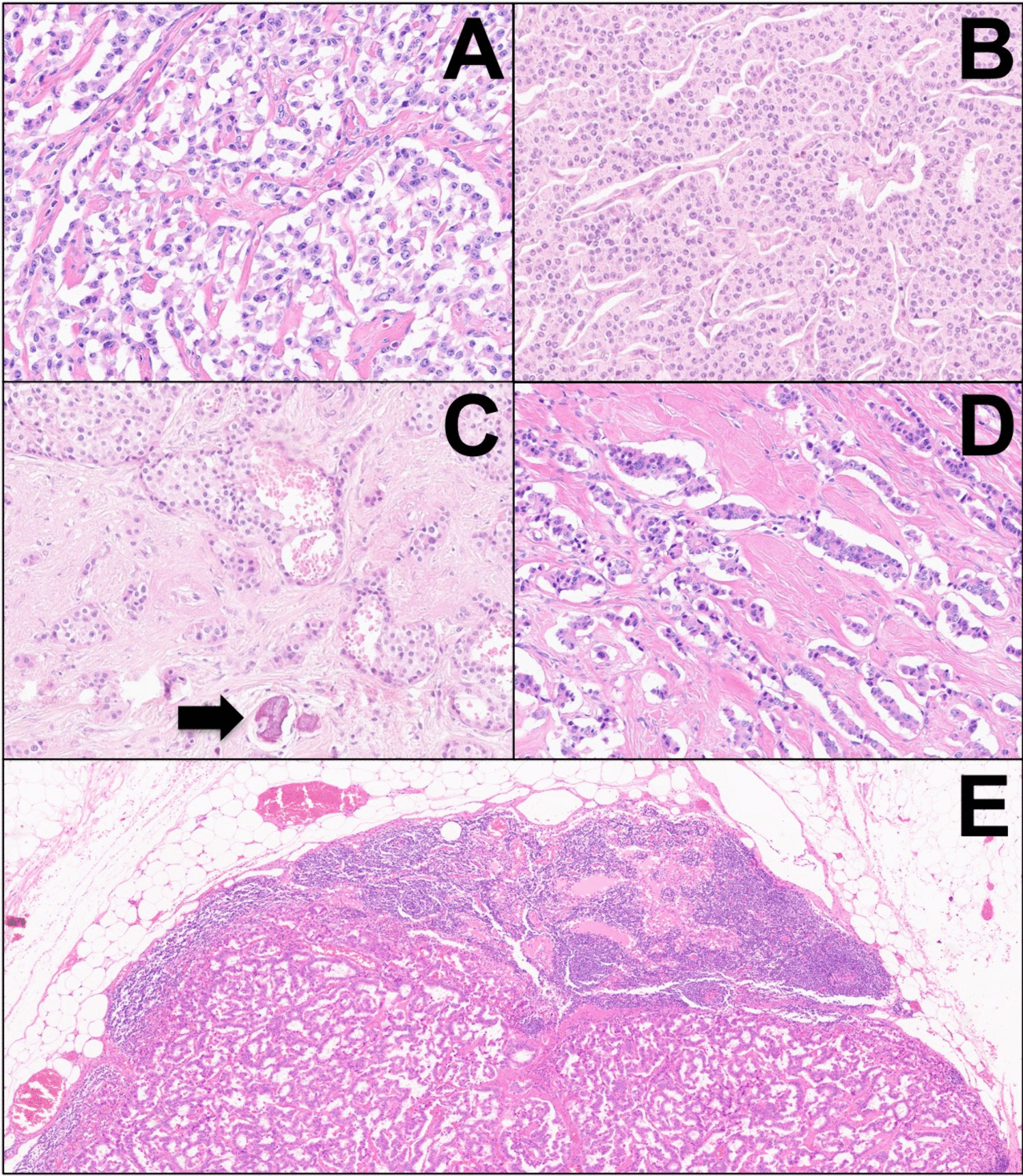
The index case was a 45-mm encapsulated angioinvasive FTC derived from a 54-year-old female patient (Fig. 1a). The diagnosis was made using the 2022 WHO classification criteria, and differentiated high-grade thyroid carcinoma could be excluded as the mitotic index was 1/2 mm2, and no tumor necrosis was evident despite extensive sampling. Venous invasion was observed multifocally, and capsular invasion was also noted. There was no extrathyroidal extension, and surgical margins were negative. Tumor cells were positive for TTF1, PAX8, and partially for thyroglobulin (50% of tumor cells). The Ki-67 index was 4.2%. Tumor cells were negative for BRAF VE1, and the P53 staining was heterogenous and therefore not indicative of an underlying TP53 gene mutation. Areas with vascular invasion were confirmed using anti-CD31 and anti-CD61 antibodies to highlight endothelial cells and thrombus material, respectively (Fig. 1b, c). The background thyroid tissue showed evidence of thyroid follicular nodular disease, but no other tumors were found. There was no sign of thyroiditis.
Fig. 1
Histological attributes of the index case. a Hematoxylin-eosin-stained section depicting extensive vascular invasion. b Double stain (CD31 in brown, pan-cytokeratin in red) visualizing intravascular tumor deposits. c CD61 immunohistochemistry confirms an in vivo phenomenon with thrombus
The clinical NGS analysis showed a TERT C250T promoter mutation. No mutations were detected in the known hotspots of various thyroid-related genes, such as the RAS gene family, BRAF, RET, MET, DICER1, PTEN, AKT, PIK3CA, APC, MTOR, and CDKN2A/B genes. No BRAF, RET, NTRK1/3, or ALK fusions were noted either. PAX8::PPARG fusions were investigated separately using PCR, with negative results.
Spatial Transcriptomics Profiling to Identify Intra-tumor Heterogeneity of Invasive Properties in FTCTo comprehensively resolve the molecular heterogeneity during the invasion process in FTCs, we performed ST on four FFPE sections (of which one showed capsular invasion, one vascular invasion, and two without invasion foci) using the 10× Genomics Visium platform (Fig. 2a, c). Transcriptomes from a total of 18,074 spots were sequenced. Detailed sequencing parameters are listed in Supplemental Table 1.
Fig. 2
Outline of the spatial transcriptomics analysis. Tiled versions of the two tissue images showing invasion that were used for spatial transcriptomics analysis are shown in a–d. a Hematoxylin-eosin-stained section of the vascular invasion area; arrows indicate the invading foci. b Unsupervised clustering analysis showing nine different clusters in slide 1 with vascular invasion. Each color represents a distinct cluster. c Hematoxylin-eosin-stained section of the capsular invasion area; arrows indicate the invading foci. d Unsupervised clustering analysis showing 10 different clusters in slide 2 with capsular invasion. Each color represents a distinct cluster (C1–10). Each spot within the 6 × 6-mm square represents an area with gene expression information, allowing for gene expression–based clustering information
Table 1 Summary of the immunohistochemical staining score of the validation cohortUnsupervised Clustering Reveals Distinct Clusters in Invading FrontsAfter identifying and annotating the morphological tumor spots, we performed an unsupervised clustering to unravel the spatial heterogeneity across the tumor. In the case with vascular invasion (slide 1), 9 different clusters were identified across the tumor, stroma, and adjacent normal tissue (Fig. 2b), while in the case with capsular invasion (slide 2), 10 different clusters were identified (Fig. 2d).
In slide 1, clusters 2 (orange) and 5 (purple) were enriched in the vascular invasion area (Fig. 2b). The top 20 DEG in cluster 2 showed significant enrichment in the GO biological process “Negative Regulation of Smooth Muscle Cell Migration” (GO:0014912) and GO cellular component “Collagen-Containing Extracellular Matrix” (GO:0062023), whereas cluster 5 showed enrichment in GO biological process “Collagen Fibril Organization” (GO:0030199) and GO cellular component “Collagen-Containing Extracellular Matrix (GO:0062023)” (Supplemental Table 2).
In slide 2, several clusters occurred both in the invasive front and central parts of the tumor; however, clusters 8 (gray) and 10 (light blue) were almost exclusively enriched in the capsular invasion area. Cluster 6 (brown) occurred in the central part of the tumor but was also intensely enriched in a separate capsular invasion area surrounded by a few spots from cluster 10 (Fig. 2d). The top 20 DEG in cluster 6 were significantly enriched in GO biological process “Extracellular Matrix Assembly” (GO:0085029). Furthermore, both clusters 8 and 10 showed enrichment in GO biological process “Extracellular Matrix Organization” (GO:0030198) (Supplemental Table 2). DEG in capsular/vascular invasion areas versus the central part of the tumor also showed enrichment in GO associated with extracellular matrix components and processes.
Among the most upregulated genes in clusters involved in the invasive areas and with biological relevance to tumorigenesis such as EMT, ECM, or tumor invasion, we selected two genes for further analysis and validation. In particular, DPYSL3 was the most upregulated gene in cluster 6 of slide 2 (Supplemental Fig. 1a–b), whereas POSTN was the most significantly up-regulated gene in cluster 10 of slide 2 and ranked in the top 5 most upregulated gene in cluster 2, slide 1 (Supplemental Fig. 1c–d).
The two areas without invasive features (slides 3 and 4) were also analyzed for differences in gene ontology, in which clones positioned near the tumoral capsule were enriched for dysregulation of genes associated to “Elastic Fiber Assembly” (GO:0048251), “Supramolecular Fiber Organization” (GO:0097435), and “Extracellular Matrix Assembly” (GO:0085029) compared to more centrally located clones (data not shown).
Furthermore, the identification of a TERT promoter mutation prompted us to investigate the expression levels of the TERT gene throughout the tumor. The analysis showed the existence of a few spots disseminated across the tumor tissue, mostly reaching both the central core and the periphery/invasive front of the tumor, suggesting an irregular expressional pattern of this gene in the index case (Supplemental Fig. 2).
Pseudotime Trajectory Analysis Reveals Angioinvasion and Epithelial-Mesenchymal Transition (EMT) as Late-Stage EventsTo gain insight into the spatial locations and to unravel spatiotemporal patterns of invasive cells, a trajectory analysis was performed on the tissue case showing angioinvasion (slide 1) after having filtered for tumor cells only by histomorphology and manual annotation of the tumor spots (Fig. 3a). The trajectory analysis revealed that tumor cells with vascular invasion clustered together and seemed to have a high pseudotime (Fig. 3b). Furthermore, after having dichotomized the tumor cells into “early” and “late” tumor cells, based on the median pseudotime (Fig. 3c, d), a gene set enrichment analysis (GSEA) was performed. In total, six gene sets were significantly differentially enriched between early and late tumor cells (Supplemental Table 3). Notably, two out of these six gene sets were associated to EMT (ECM-receptor interaction and cell adhesion molecules) (Fig. 3e, f). Both gene sets were negatively enriched in early versus late tumor cells.
Fig. 3
Trajectory analysis and gene set enrichment analysis in the angioinvasive tissue sample. a Hematoxylin-eosin-stained section of angioinvasive tissue with filtered tumor cell spots for analysis. b Trajectory analysis with tumor cell spots, showing clustering of angioinvasive tumor cell spots occurring with high pseudotime. c Hematoxylin-eosin-stained section of angioinvasive tissue representing the time stage of “early” and “late” tumor cells. d Trajectory analysis dichotomized into “early” and “late” tumor cells based on median pseudotime. e Gene Set Enrichment Analysis (GSEA) plots demonstrating downregulation of ECM-receptor interaction (path:hsa04512) in early versus late (NES −2.03, p-value 0.00, FDR 0.00). f Gene set enrichment analysis (GSEA) plots showing the downregulation of cell adhesion molecules (path:hsa04514) in early versus late (NES −0.51, p-value 0.00, FDR 0.05)
Distinct Expression Gradient of Extracellular Matrix (ECM)–Related Markers from Tumor Center to PeripheryTwo of the DEG in the invasive clusters were selected to verify and validate our initial observations shown by ST analysis. Immunohistochemical staining for cytoplasmic proteins DPYSL3 and POSTN was performed in an independent cohort of 20 FTCs and 10 FTAs.
The staining revealed an exclusive cytoplasmic localization for both markers. The level of immunoreactivity of both proteins in FTAs was homogeneous throughout the tumor, both in the central core and periphery of the tumor. Among the 10 FTAs stained for POSTN, 8 cases received a score of 1 in the central part of the tumor, while 9 were scored 1 in the periphery. Similar results were obtained with DPYSL3: 6 cases scored 1 in the central part of the tumor, while 9 obtained a score of 1 in the periphery. None of the 10 FTAs stained for both markers received a score of 2 (Table 1). No significant differences were found in comparing the central part versus the periphery of the tumor for both proteins (Fig. 4a, b).
Fig. 4
Extracellular matrix (ECM) marker immunohistochemistry. Immunohistochemical staining of DPYSL3 and POSTN in follicular thyroid carcinoma (FTC) compared to follicular thyroid adenoma (FTA). a, b Staining showing the homogeneous expression of DPYSL3 and POSTN respectively in FTA. c, d Pronounced expression of DPYSL3 and POSTN, respectively, characterized by increased intensity extending from the central region towards the periphery of the FTC. e, f Strong expression of DPYSL3 and POSTN respectively in the area with capsular invasion. Statistical significance for this feature was observed only for DPYSL3. g, h Pronounced expression of DPYSL3 and POSTN in the area of vascular invasion. Statistical significance for this feature was observed only for DPYSL3
Among the 20 FTCs stained for DPYSL3, 12 cases (60%) exhibited central tumor staining (score 1). In contrast, all 20 cases (100%) stained for DPYSL3 displayed a distinct expression pattern, characterized by heightened intensity extending from the central region of the tumor towards the periphery, particularly in proximity to the tumor capsule. The same expression pattern was observed in FTCs stained for POSTN, where 11 cases (55%) showed central tumor staining (score 1), and 19 cases (95%) exhibited moderate to strong positivity, particularly near the tumor capsule (Table 1; Fig. 4c, d). Significant differences were noted when comparing the immunoreactivity levels of the central core part versus the periphery of the tumor (p = 0.001) for both DPYSL3 and POSTN proteins. Moreover, strong cytoplasmic staining was also evident in areas exhibiting capsular (Fig. 4e, f) and vascular invasion (Fig. 4g, h). All 20 cases (100%) stained for DPYSL3 showed moderate or strong expression intensity around the capsular or vascular invasion focus, showing a significant difference compared to a more central part of the tumor (p = 0.002). Sixteen out of 20 FTCs (80%) stained for POSTN exhibited moderate or strong expression intensity around the capsular or vascular invasion focus; however, no significant differences were found in comparing the central core versus the invasive front of the tumor.
Comments (0)