Glutathione S-transferases (GSTs) are widely distributed in nature from bacteria and yeasts to plants and animals (Mannervik et al., 2005; Allocati et al., 2009). They play a key role in enzymatic detoxification by catalysing the nucleophilic addition of the thiol group of glutathione to a wide variety of electrophilic endo- and xeno-substrates (Armstrong, 1997; Hayes and Pulford, 1995). In addition, GSTs play a role in several cellular processes unrelated to detoxification. These include biosynthesis of sex steroids and prostaglandins (Johansson and Mannervik, 2001; Kanaoka et al., 1997), JNK signaling (Adler et al., 1999), the MAP kinase pathway (Cho et al., 2001), increased risk of numerous human diseases, including cardiovascular disease (Maciel et al., 2009), Parkinson's and Alzheimer's diseases (Li et al., 2003) and a new immuno-regulatory strategy against inflammatory bowel disease (Driss et al., 2016; Sarazin et al., 2018; Capron et al., 2019; Foligné et al., 2019).
Schistosomes are parasitic worms responsible for a human infection termed schistosomiasis, which is considered by the World Health Organization as the second most devastating parasitic infection after malaria (Lo and Arnold, 2023). Schistosomes have two main GST isoforms, namely, 26 kDa and 28 kDa, which differ in substrate specificities (Walker et al., 1993). Currently, it is not known which isoform is necessary for parasite survival. The SjGST-26 GST isoform is also well-known in biological laboratories, being one of the commercial plasmids used for GST fusion protein expression (Smith et al., 1998; Kim and Hakoshima, 2019; Gnanasekaran and Pappu, 2023).
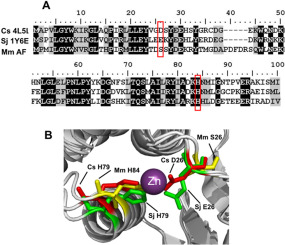
Earlier reports did not show a natural association of GST with metal ions (Chaga et al., 1994); however, Yoshinaga et al. (2006)] identified GSTs capable of binding vanadium in the ascidian Ascidia sydneiensis samea. In Clonorchis sinensis, the crystal structures of CsGST in two different spatial groups revealed that Asp26 and His79 coordinate a zinc ion (Han et al., 2013). In homologous positions, Glu26 and His79 seem to be responsible for the weak Ni2+-binding affinity of SjGST-26 in Immobilized Metal Affinity Chromatography (IMAC) (Han et al., 2010). In concert with this, enhancement of SjGST-26 Ni2+-binding affinity is observed following mutation of Glu26 to histidine. Except for potential involvement in metal homeostasis and metal detoxification, the role of these zinc-binding sites has not been previously addressed.
The SLC6 family of sodium/chloride-dependent neurotransmitter transporters are membrane proteins containing 12 transmembrane helices and a large extracellular glycosylated loop, with the N- and C-termini oriented into the cytoplasm (Kristensen et al., 2011). In some of them, namely dopamine transporter (DAT) (Norregaard et al., 1998), glycine transporter 1 (GlyT1) (Ju et al., 2004) and mouse GABA transporter 4 (GAT4) (Cohen-Kfir et al., 2005), (the equivalent of rat GAT3), Zn 2+-binding sites have been identified. These zinc-binding sites likely have a physiological function, since in brain zinc acts as the neurotransmitter and neuromodulator (Norgaard-Nielsen and Gether, 2006). This function of zinc is supported by the existence of zinc-containing synaptic vesicles, filled by a specific zinc transporter ZnT3 (also named Slc30a3) (Palmiter et al., 1996; Wenzel et al., 1997). Thus, zinc can be released on nearby zinc binding receptors and transporters to modulate their function (McMahon and Cousins, 1998; Palmiter and Huang, 2004).
In this work, we show that the manganese/zinc binding sites of both SjGST-26 and neurotransmitter transporters GlyT1 and GAT3 can coordinate their mutual interaction in vitro. This suggests that these metal binding sites might serve as coordination sites for their interaction with other metalloproteins in vivo. This is especially interesting because both SjGST-26 and zinc were previously shown to play important roles in schistosome infections (Brophy and Pritchard, 1994; Asch and Dresden, 1977; Friis et al., 1997). SjGST-26-tagged proteins are frequently used in protein pull-down assays (abbreviated as GST pull-assays) (Kim and Hakoshima, 2019; Gnanasekaran and Pappu, 2023). Our work indicates that when such an assay involves metalloprotein targets, the presence of metal ions may lead to false-positive interactions.


留言 (0)