
記住我
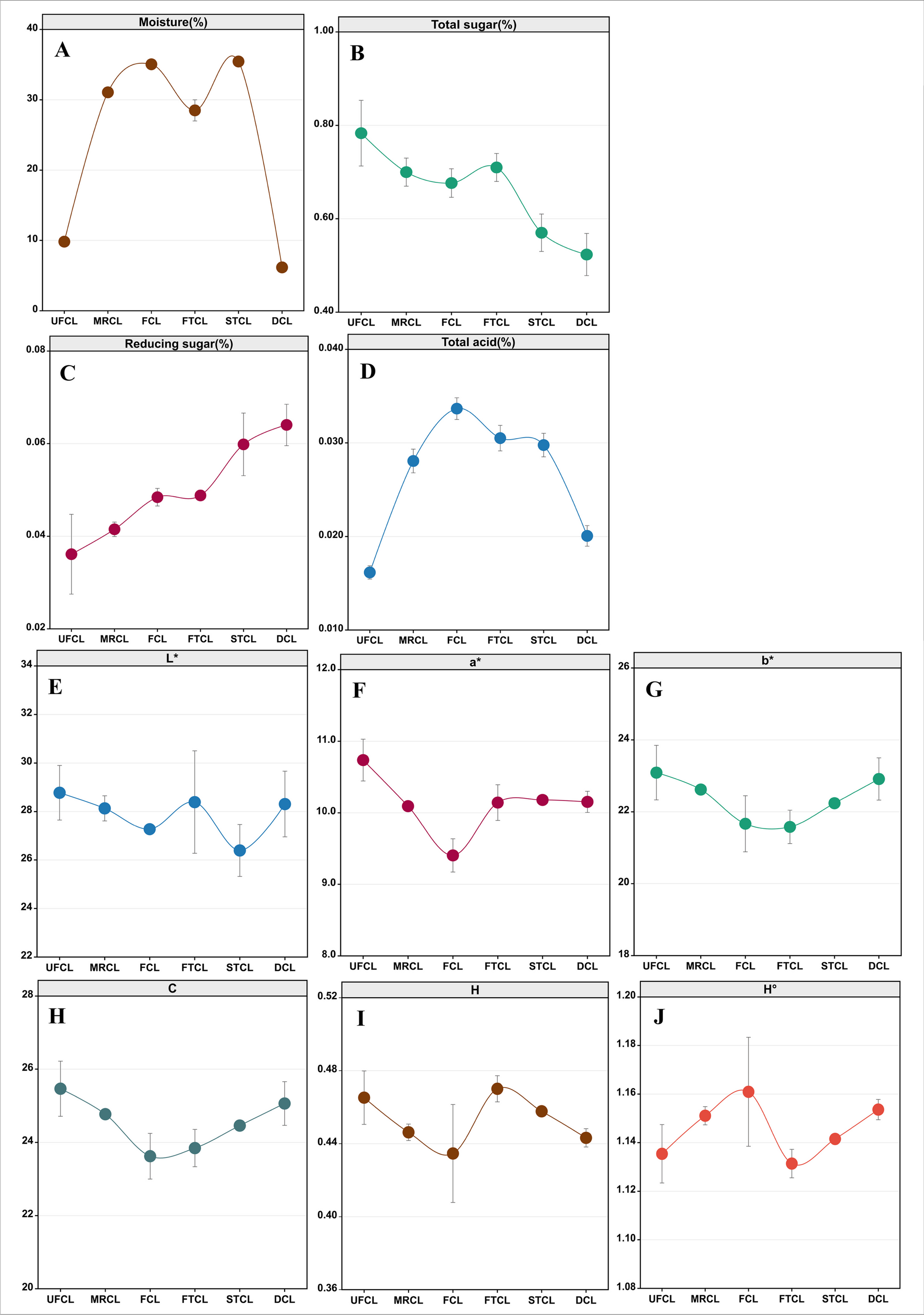

During the CTL fermentation process, we investigated the dynamic changes in moisture content, chroma, total sugar, reducing sugar concentration, and total acidity (Fig. 1). After rehydration, the moisture content of CTLs increased fast, with the moisture content of UFCL to MRCL increasing from 9.82 to 31.06%. As the fermentation time progressed, a decreasing trend in total sugar content and a gradual increase in reducing sugar content were observed. The reduction in chemical component concentration in tobacco leaves is attributed to enzyme activity and microbial metabolism. Specifically, during the fermentation process, activities such as amylase increase, leading to starch hydrolysis and the formation of reducing sugars, as demonstrated in previous studies (Yamaguchi et al. 2013). The total sugar concentration, on the other hand, does not grow, possibly because bacteria use these sugars as resources to support their metabolism. In contrast, the total acidity of CTLs was initially low in the UFCL samples at the beginning of fermentation and then increased before drying. The total color difference (▲E*) represents the overall change in color during agricultural product processing or storage, incorporating the three fundamental color parameters: L*, a*, and b*. A higher value indicates a more pronounced color change. The trends of the tobacco leaf color parameters L*, a*, b*, C, H, and H° during the fermentation process are depicted in Fig. 1. It can be observed that there are significant differences in the tobacco leaf color parameters during fermentation. With the progression of fermentation, the L*, a*, b*, and C values initially decrease and then increase, with the most substantial changes occurring after the first turning of the fermentation pile. Subsequently, the magnitude of the changes gradually diminishes, reaching a stable state. As the fermentation progresses, the H value follows a pattern of initial decrease, subsequent increase, and final decrease. The highest value is observed during the first turning of FTCL, followed by a gradual decline. Conversely, the H° value demonstrates an initial increase, followed by a decrease, and then another increase. The lowest value is observed during the first turning of FTCL, after which there is a gradual rise.
Fig. 1
Changes in the physicochemical indices during different stages of fermentation for CTLs. A Moisture; B total sugar; C reducing sugar; D total acid; and chromatic parameters E L*, F a*, G b*, H C, I H, and J H°. The stages of fermentation are denoted as UFCL (unfermented cigar leaves) representing leaves before moisture reacquisition, MRCL (moisture-regained cigar leaves) indicating leaves post moisture reacquisition, FCL (fermented cigar leaves) denoting leaves subsequent to moisture reacquisition and fermentation, FTCL (first turn cigar leaves) representing leaves after the initial turn, STCL (second turn cigar leaves) indicating leaves post second turn, and DCL (dried cigar leaves) representing leaves after the completion of fermentation and drying
We performed regression analysis to examine the relationship between the physicochemical parameters of CTLs during industrial fermentation and the total color difference (▲E*). Using moisture content, total sugar, reducing sugar, and total acidity as dependent variables and ▲E* as the independent variable, we established regression equations for each physicochemical indicator at different fermentation stages (Figure S3). The ▲E* value exhibited a gradual upward trend (Figure S2), indicating a progressive darkening of the color in CTLs during fermentation. The regression analysis revealed that moisture content, reducing sugar, and total acidity had a direct and positive impact on the ▲E* value, while total sugar content had a direct and negative impact. In summary, the physicochemical parameters directly influenced the color changes of CTLs throughout the fermentation process.
Analysis of volatile compoundsThe profiles of volatile compounds (VOCs) in CTLs were detected using GC–MS to explore the impact of CTLs fermentation on volatile flavors. Eight aldehydes, 25 ketones, 3 esters, 7 alkanes, 4 terpenes, 2 nicotinoids, 5 alcohols, 2 phenols, and 7 additional volatile scents were found during fermentation. In all CTLs samples, the bulk of volatile chemicals (39.68% of all volatile components) were ketones (Figure S4).
The heatmap analysis results show that the metabolic profiles of fermented CTLs have various stages, as shown in Fig. 2A. In phases 1 and 3, there was a notable increase in both the diversity and concentration of metabolites compared to phase 2. Among these, ketone compounds were particularly variable. Out of the 25 ketone compounds identified, 16 flavor components showed significant fluctuations throughout these phases. The three VOCs with the most marked variability were 4,7,9-megastigmatrien-3-one, geranylacetone, and solanone. Their relative concentrations varied significantly across the three phases, with ranges of 77.50 to 126.35 µg/µL for 4,7,9-megastigmatrien-3-one, 40.83 to 77.81 µg/µL for geranylacetone, and 49.16 to 85.83 µg/µL for solanone. Besides, volatile nicotine and neoplantene emerge as the predominant compounds in CTLs, constituting 77.81 to 94.76% of the total volatiles. Throughout the fermentation process, the relative content of volatile nicotine initially rises, then declines (50.87% vs 80.12% vs 51.98%), whereas neoplantene exhibits a decrease followed by an increase (26.94% vs 11.35% vs 28.56%). This fluctuation in volatile nicotine may be attributed to the conversion of alkaloids in CTLs and their subsequent release into the air during deep fermentation. Neoplantene, a degradation product of chlorophyll, itself lacks a strong aroma or fragrance but significantly influences sensory attributes such as aroma quality, smoothness, and transmittance.
Fig. 2
A Heatmap analysis was performed to examine the volatile compounds in CTLs during fermentation; B PLS-DA score plots of volatile flavors were generated to analyze their characteristics; C permutation tests were performed to compare the three different stages of fermentation, and D key volatile compounds were identified using PLS-DA, with a VIP score of > 1.0 and an OAV score of ≥ 1.0, indicating their significance and contribution. CTLs are classified into three phases: phase 1 (before moisture regaining, including UFCL), phase 2 (after moisture regaining, including MRCL, FCL, FTCL, and STCL), and phase 3 (after fermentation and drying, including DCL)
Based on the analysis of 63 VOCs as the dependent variable and different fermentation stages as the independent variable, the results of PLS-DA (Fig. 2B) and PCA (Figure S4) demonstrate effective discrimination of CTLs at various fermentation stages. PLS-DA is a supervised multivariate statistical analysis technique that combines metabolite changes with experimental grouping information through regression models while reducing dimensions. To assess model’s quality, we utilize metrics such as R2X, R2Y, and Q2. The goodness of fit indices for the independent variables, represented by R2x (0.724), and the dependent variables, represented by R2y (0.961), indicate satisfactory model fitting. Additionally, model’s prediction index, Q2 (0.926), surpasses the threshold of 0.5, further supporting the reliability of the model (Yun et al. 2021). The validity of the model is confirmed by performing 200 permutation tests, as depicted in Fig. 2C, where the intersection between the Q2 regression line and the y-axis is negative, indicating the absence of overfitting. As a result, these data can be used to identify and analyze different fermentation phases in CTLs. Furthermore, a VIP score threshold of > 1.0 (Figure S4) was employed in combination with the screening criterion of olfactory activity value (OAV) > 1.0 (Table S3) to assess the impact strength and explanatory power of differential metabolite accumulation differences on the classification and discrimination of samples in each group. In total, 15 metabolites exhibited substantial differences among the three groups and made significant contributions to the overall aroma of CTLs. These metabolites are beta-cyclocitral, decyl aldehyde, phenylacetaldehyde, acetophenone, isophorone, 2,6,6-trimethyl-2-cyclohexene-1,4-dione, 1-(3,4,5-trimethylphenyl) ethanone, farnesylacetone, tetrahydro-actinidiolide, undecane, (-)-alpha-terpineol, 2,6-di-tert-butyl-4-methylphenol, 3-acetylpyridine, quinoline, and indole. Figure 2D depicts the temporal fluctuations of these 15 metabolites during the fermentation process. In Fig. 2D, distinctive circular shapes of various colors represent 15 different types of differential metabolites, while the unfilled circles denote volatile metabolites that were not detected in this specific sample. It is readily apparent that 11 and 13 differential metabolites were identified in UFCL and DCL, respectively, providing further evidence of the higher diversity in both types and quantities of metabolites during phase 1 and phase 3 in comparison to phase 2. Notably, among the detected differential metabolites, ketones constitute 45.45% and 38.46% in UFCL and DCL, respectively, showcasing a more prominent variability.
Microbial community composition diversity and differencesThe researchers observed the industrial fermentation process of CTLs to evaluate the microbial community makeup among different groups. Following the application of quality control procedures, 18 samples yielded 3,042,137 high-quality sequences that represented communities of bacteria. The number of sequences in the sample ranged from 74,366 to 219,738. Also obtained were 2,300,041 high-quality reads from fungi, with read counts per sample ranging from 111,954 to 136,222. The sequence numbers retrieved were sufficient to reach saturation, according to the goods coverage analysis (Table S2).
At the phylum level, the bacterial community was dominated by members of the Firmicutes phylum, which accounted for 94.73% of the relative abundance at the final stage of fermentation (Fig. 3A). Moving down to the genus level, during the initial stages of fermentation (UFCL/Phase1), Corynebacterium, Pseudomonas, Staphylococcus, and Stenotrophomonas displayed higher abundance. However, following the rehydration of CTLs, Aerococcus emerged as the dominant genus and maintained its prevalence throughout the fermentation process (Fig. 3B). Shifting the focus to the fungal aspect, the Ascomycota phylum dominated the fungal population at the start of fermentation, accounting for 96.82% of the total microbial abundance (Fig. 3C). At the genus level (Fig. 3D), prior to and after moisture regaining (UFCL/MRCL), Aspergillus exhibited dominance, with relative abundances of 50.68% and 64.44% in UFCL and MRCL, respectively. Alternaria and Cladosporium were the dominant genera in the accumulated CTLs pile, accounting for relative abundances of 35.28% and 31.96%, respectively. Notably, a slight increase in fungal diversity was observed in the FTCL during fermentation, with Aspergillus, Alternaria, and Cladosporium emerging as the dominant genera, comprising abundances of 34.22%, 24.90%, and 28.86%, respectively. Aspergillus maintained dominance in both STCL and DCL, with abundances of 62.01% and 55.73%, respectively.
Fig. 3
The distribution of the microbial community in CTLs at various stages of fermentation. Bacteria (A) and fungus (C) at the phylum level; bacteria (B) and fungi (D) at the genus level
By analyzing α-diversity and β-diversity, microbial richness and diversity can be calculated. The fermentation process can be separated into stages 1, 2, and 3 based on the results of the diversity and cluster analysis. Shannon and Chao index values of CTLs showed a relatively high level of diversity in phase 1 (pile fermentation: CTLs before moisture recovering). In phase 2 (pile fermentation: CTLs after moisture regaining, fermentation, and turning), Shannon’s diversity index and Chao index values initially decreased and subsequently either increased or remained relatively stable. In phase 3 (CTLs after fermentation and drying), Shannon’s diversity index and Chao index values of CTLs all decreased to a comparatively lower level. Moreover, to explore the divergent microbial communities, we conducted β-diversity analysis for the three groups. As depicted in Fig. 4A–B, the principal coordinates analysis based on the Bray–Curtis distance matrix demonstrated the distinct separation of bacterial and fungal communities into three distinct clusters. The different fermentation processes had specific impacts on both bacterial and fungal populations. However, when compared to the fungal PCoA distribution trend (Fig. 4B), the differentiation of bacterial microbiota at different stages was more pronounced (Fig. 4A). These disparities in bacterial microbial community composition might be more substantial, indicating that bacteria could be pivotal in the dynamic changes observed in the metabolite profile during fermentation stages.
Fig. 4
The dynamics of the microbial community in CTLs at various stages of fermentation. The following are the results of principal coordinate analysis (PCoA) of bacterial (A) and fungal (B) communities using the Bray–Curtis distance matrix; indicator analysis; and cross-validation statistical tests: bacteria (C) and fungi (D)
Indicator value analysis was conducted to compare the species richness across the three stages of CTLs and identify the significant differences in the studied genera. A comprehensive assessment of bacterial genera revealed a total of 13 genera that exhibited noteworthy significance as indicators for distinct fermentation stages. Similarly, for fungal genera, a total of 7 genera demonstrated significant indicator values (indicator value p < 0.01). Remarkably, the microbial genus assemblages indicative of phase 1, 2, and 3 fermentation stages, as classified based on CTLs’ diverse industrial fermentation processes, remained consistently present throughout the sampling period. However, as quantified by the coefficient of variation, certain time points displayed higher relative abundances and lower variation (Fig. 4C–D).
Relationship between the microbial community, physicochemical characteristics, and volatile compoundsIn all CTLs samples, Pearson correlation analysis was carried out (R > 0.5, p < 0.05; Fig. 5A–B) to investigate any potential relationships between the dominating genera and four important environmental parameters (moisture, total acid, total sugar, and reducing sugar content). The results unveiled intriguing associations between these factors and the microbial composition. Notably, a positive correlation was observed between total sugar and bacterial genera. Conversely, moisture and total acid displayed negative correlations with all bacterial genera, except for Facklamia (Fig. 5A). It is important to note that fungi were affected by environmental conditions differently than bacteria were. Both total sugar and reducing sugar exhibited positive correlations with five selected fungal genera, while moisture and total acid displayed negative correlations with all genera, with the exception of Xeromyces (Fig. 5B). These correlation analyses may offer a new insight into the complex dynamics of CTLs fermentation by highlighting the complicated interplay between physicochemical characteristics and microbial communities.
Fig. 5
The correlation network between selected bacteria and physicochemical indicators (A), and the correlation network between selected fungi and physicochemical indicators (B). Pearson’s correlation coefficient (R > 0.5, p < 0.05) was used to calculate the selected bacteria or fungi; variation partitioning analysis of the relative contributions of moisture, reducing sugar, total sugar, and total acid to variation in CTLs bacterial β-diversity (C); and variation partitioning analysis of the relative contributions of moisture, reducing sugar, total sugar, and total acid to variation in CTLs fungal β-diversity (D). Heat map of correlation between bacterial (E) with fungal (F) and VOCs; blue and green represent positive correlation, red and purple represent negative correlation, and the size of the circles represents the magnitude of the |R| value; significant correlations were expressed by *, **, ***, and ****, which were p < 0.05, p < 0.01, p < 0.001, and p < 0.0001, respectively. VOCs2, beta-cyclocitral; VOCs3, decyl aldehyde; VOCs7, phenylacetaldehyde; VOCs10, acetophenone; VOCs11, isophorone; VOCs12, 2,6,6-trimethyl-2-cyclohexene-1,4-dione; VOCs26, 1-(3,4,5-trimethylphenyl) ethenone; VOCs28, farnesylacetone; VOCs36, tetrahydro-actinidiolide; VOCs39, undecane; VOCs53, (-)-alpha-terpineol; VOCs55, 2,6-di-tert-butyl-4-methylphenol; VOCs58, 3-acetylpyridine; VOCs62, quinoline and VOCs63, indole
The impact of moisture, reducing sugar, total sugar, and total acid on microbial community variation is illustrated in a bar diagram (Fig. 5C–D). The comprehensive analysis of all variables accounted for 61.87% and 8.51% of the variation in the bacterial and fungal communities of CTLs, respectively. Among the bacterial community, the major contributor was total sugar, which accounted for 30.71% of the variation. Additionally, total acid (21.94%) made a substantial contribution, while moisture (5.11%) and reducing sugar (4.11%) had comparatively smaller effects on bacterial β-diversity in CTLs. Regarding the fungal community, total acid (3.39%) emerged as the primary contributing factor. On the other hand, moisture (− 1.47%), reducing sugar (− 1.62%), and total sugar (− 2.03%) displayed negative contributions to the fungal community variation.
To explore the intricate relationship between bacteria and flavor development during fermentation, we conducted a comprehensive analysis using a correlation heat map and cluster analysis. The correlations between bacterial and fungal communities and the metabolite profiles of fermented CTLs may be divided into three different groups, according to our findings, which are shown in the hierarchical cluster analysis (HCA) results (Fig. 5E–F). In terms of bacterial microorganisms, cluster 1 consisted of VOCs10 (acetophenone), VOCs3 (decyl aldehyde), VOCs53 ((-)-alpha-terpineol), and VOCs2 (beta-cyclocitral). These compounds exhibited significant positive correlations with Pseudomonas, Staphylococcus, Stenotrophomonas, Flavobacterium, Sphingomonas, Allorhizobium-Neorhizobium-Parararhizobium-Rhizobium, Sphingobium, and Agrococcus were also significantly positively correlated while showing significant negative correlations with Aerococcus. Cluster 2 consisted of VOCs39 (undecane), VOCs11 (isophorone), VOCs28 (farnesylacetone), VOCs12 (2,6,6-trimethyl-2-cyclohexene-1,4-dione), VOCs58 (3-acetylpyridine), and VOCs26 (1-(3,4,5-trimethylphenyl) ethenone). These compounds displayed significant positive correlations with the nine bacterial genera mentioned earlier, although with smaller R-values and weaker correlations compared to cluster 1. Cluster 3, consisting of VOCs63 (indole), VOCs7 (phenylacetaldehyde), VOCs55 (2,6-di-tert-butyl-4-methylphenol), VOCs62 (quinoline), and VOCs36 (tetrahydro-actinidiolide), exhibited a distinct pattern. Notably, cluster 3 showed a significant negative correlation with Corynebacterium and a significant positive correlation with Bacteroides, distinguishing it from the other clusters. Regarding fungal microorganisms, cluster 1 encompassed VOCs10, VOCs3, VOCs28, and VOCs12, which demonstrated significant positive correlations with Colletotrichum, Cumuliphoma, and Fusarium. Cluster 2 comprised VOCs58, VOCs11, VOCs26, VOCs39, VOCs53, and VOCs2. These compounds exhibited some positive correlations with the aforementioned three fungal genera, with VOCs53 and VOCs2 showing significant positive correlations with Nicotiana, Symmetrospora, Septoria, Golubevia, and Moesziomyces. Cluster 3, consisting of VOCs63, VOCs7, VOCs55, VOCs62, and VOCs36, displayed distinct associations. Notably, the clustering pattern in cluster 3 for bacterial genera was consistent, suggesting a consistent trend in the effects of these VOCs on the microflora. Cluster 3 exhibited a significant positive correlation with Sterigmatomyces and a significant negative correlation with Rhodotorula.
Metabolic map of important volatile compounds during the industrial fermentation of cigar tobacco leavesTo gain deeper insights into the association between microorganisms and VOCs, we employed the Kyoto Encyclopedia of Genes and Genomes (KEGG) database and relevant literature to predict the metabolic network of significant VOCs during the industrial fermentation of CTLs. Analysis of the metabolic pathways using the KEGG and MetaCyc databases revealed that the synthesis and biosynthesis pathways of key flavor compounds were generally consistent. Figure 6 illustrates the impact of the bacterial community on the profiles of VOCs through the modulation of relevant enzyme activities.
Fig. 6
Metabolic map of important VOCs during the industrial fermentation of CTLs, A the pathway of carotenoid biosynthesis and degradation, B aromatic amino acid biosynthesis leading to Trp, Tyr, and Phe in plants, and C biosynthetic and degradation pathway of cembranoid diterpenes. The MEP pathway is represented by the pathway in the rectangle above, which produces substrates for carotenoid synthesis. The pathway in the bottom ellipse illustrates carotenoids’ synthesis and degradation in plastids. Enzymes are depicted in blue
The VOCs present in CTLs can be categorized into distinct groups based on their origin, including benzene carotenoid degradation products, phenylalanine degradation products, cypress-like substances, and chlorophyll degradation products (neophytadiene). As depicted in Fig. 6A, carotenoids are synthesized through the condensation of isopentenyl diphosphate (IPP) and dimethylallyl diphosphate (DMAPP), which are generated via the 2-C-methyl-d-erythritol 4-phosphate pathway (MEP) (Klee and Tieman 2013). Crucial enzymes involved in this pathway, such as EC: 2.2.1.7 (1-deoxy-d-xylulose-5-phosphate synthase) and EC: 1.1.1.267 (1-deoxy-d-xylulose 5-phosphate reductoisomerase), play a fundamental role in determining the production rate of IPP. It is important to note that the content and composition of carotenoids in plants are not solely influenced by the biosynthesis pathway but also by the degradation pathway. Carotenoids serve as essential precursors for various bioactive compounds. They can undergo enzymatic cleavage, primarily catalyzed by EC: 1.13.11.68 (carotenoid cleavage dioxygenases) and EC: 1.13.11.51 (9-cis-epoxycarotenoid dioxygenases), or non-enzymatic reactions initiated by reactive oxygen species (ROS). Carotenoid degradation products, such as β-ionone, geranylacetone, farnesylacetone, and megastigmatrienone, are significant VOCs found in CTLs. Enhancing the levels of VOCs through increased carotenoid degradation may be considered an effective approach due to the positive association between carotenoid degradation and flavor formation.
Phenylalanine (Phe) and tryptophan (Trp) are considered essential aromatic amino acids, whereas tyrosine (Tyr) is deemed non-essential (Fürst and Stehle 2004). The synthesis of these three amino acids begins with the shikimate pathway, which converts phosphoenolpyruvate and erythrose 4-phosphate into chorismate, which is then converted into Phe and Trp via the aromatic amino acid biosynthetic pathways (Tzin and Galili 2010) (Fig. 6B). Notably, the metabolic pathway of phenylalanine involves the formation of phenylacetic acid (Yang et al. 2011) and phenylacetaldehyde, which can be linked to specific enzymes to establish a comprehensive metabolic pathway. This pathway can be divided into two primary directions: the first involves the conversion of phenylalanine into phenylglyoxal in the presence of EC: 4.1.1.28 (aromatic-l-amino-acid decarboxylase), followed by the production of phenylacetaldehyde through the action of EC: 1.4.3.21 (primary-amine oxidase). The second direction involves the formation of phenylalanine into 2-phenylethylamide in the presence of EC: 1.11.1.21 (catalase-peroxidase), followed by the conversion to phenylacetic acid by EC: 3.5.1.4 (amidase). Floral and fruity aromas in CTLs primarily originate from esters and aromatic compounds. Aromatic compounds are characterized by floral, fruity, and sweet scents, with distinct aromas, high boiling points, and limited volatility, making them crucial contributors to the flavor composition of CTLs. Phenylglyoxal, found in the phenylalanine metabolic pathway, is a significant aromatic flavor compound in CTLs, contributing floral and fruity aromas. Consequently, the metabolism of phenylalanine in CTLs can generate floral and fruity aromas in cigars.
Cembranoid diterpenes, commonly found in plants, are synthesized from geranylgeranyl diphosphate (GGPP) through the catalytic action of cembratrienol synthase (CBTS) and cytochrome P450 hydroxylase (CYP450) in the 1-deoxy-d-xylulose-5-phosphate pathway (DXP). GGPP, an intermediate in the DXP pathway of plant terpene metabolism, is synthesized from IPP and DMAPP (Fig. 6C). In tobacco, the biosynthesis of cyproterane diterpenes occurs in two steps: firstly, GGPP provides the cyproterane skeleton structure for the formation of cembratrienol (CBT-ol), which undergoes cyclization to form α- and β-CBT-ol in the presence of CBTS (Guo and Wagner 1995); secondly, through the action of CYP450, the sixth carbon of CBT-ol can be degraded to produce various aroma components such as ketones and their derivatives during the growth and processing of tobacco leaves (Wang et al. 2001). Ketones themselves possess a pleasant aroma reminiscent of carrots, licorice, and tea. With further modification and aging of tobacco, ketones can undergo degradation to ketofurans and ketone oxides, which significantly contribute to the flavor of cigarettes. Ketone oxides, for example, impart a subtle nutty and sweet aroma, enhancing the overall aroma profile of cigarettes with notes of cocoa and mild tobacco.
留言 (0)