
記住我
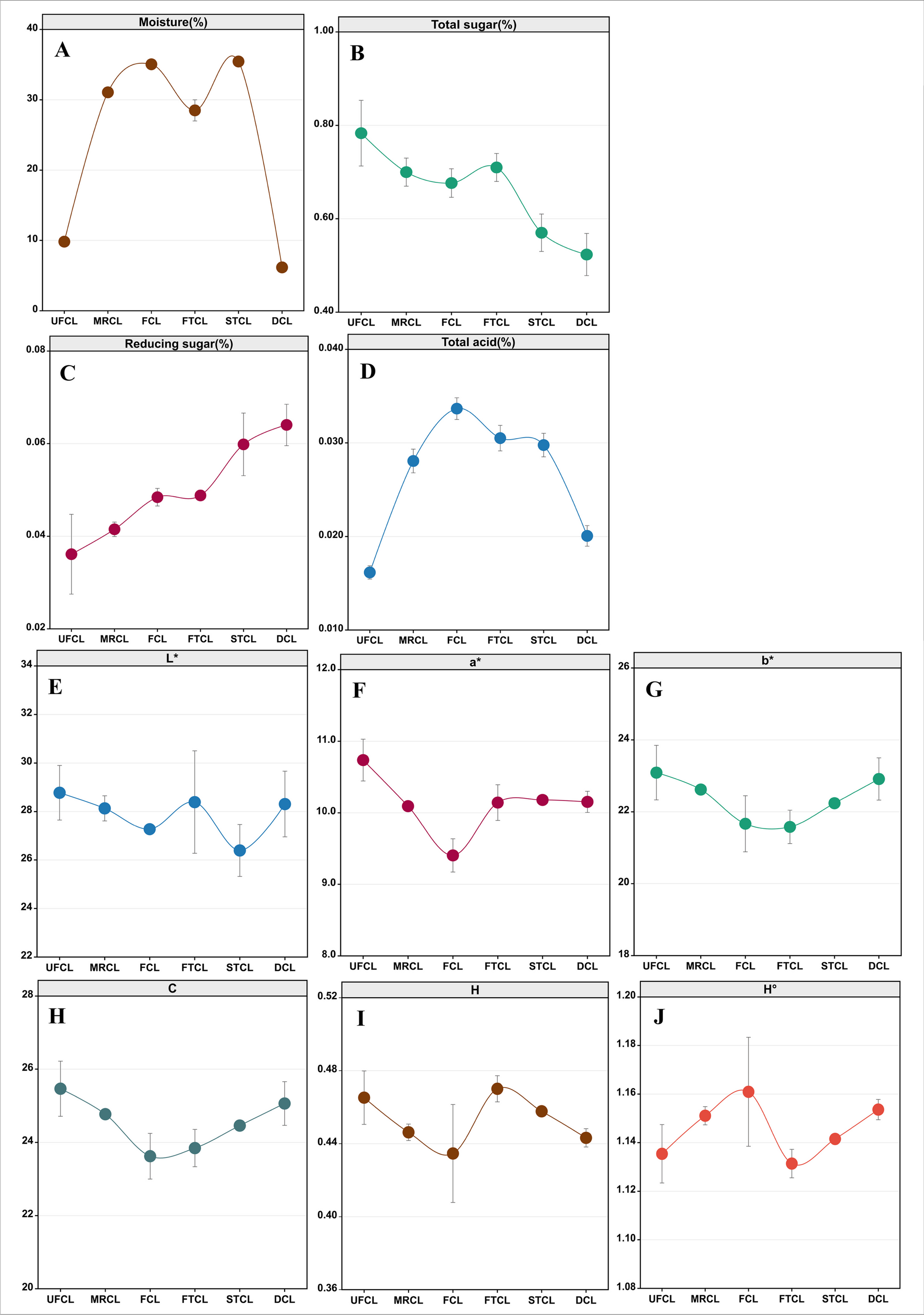

To obtain strains that were able to grow at high temperatures, ALE and mutagenesis were adopted using erythritol-high-producing Y. lipolytica CA20 as a start (Liu et al. 2023). The breeding process was divided into four stages, with temperatures ranging from 30 to 34 °C, 34 to 36 °C, 36 to 37 °C, and 37 to 38.5 °C, respectively (Fig. 1). In each stage, diverse methods were applied. To avoid the loss of erythritol synthesis by thermotolerant strains, glucose of 200 g/L was added to the medium to maintain the osmotic stress, and TTC was also added as an indicator (Qiu et al. 2021; Liu et al. 2023). The results showed that it took 150 days (corresponding to 352 generations) to obtain a strain capable of growing at 38.5 °C, during which the time needed for each stage was 10 (16 generations), 20 (42 generations), 30 (69 generations), and 90 (225 generations) days, respectively. The cell shape of CA20 grown at 30 °C was oval, while the shape of HT385 was elongated and oblate when grown at 38.5 °C, indicating a physiological alteration of the cell during adaptive evolution.
To determine whether the obtained thermotolerant strains were stable or just a tentative phenotype change, we next detected the growth of Y. lipolytica HT34, Y. lipolytica HT36, and Y. lipolytica HT385 under different temperatures. As shown in Fig. 2, the growth of HT34 at 30 °C was similar to that observed for CA20, whereas the strains HT36 and HT385 grew better than HT34 and CA20, especially for HT385 where the maximum OD600 was close to 25 after growing for 96 h (Fig. 2a). CA20 was not able to grow at 34 °C, while the strains HT34, HT36, and HT385 grew well (Fig. 2b). The thermotolerant phenotype of HT34 was stable, as there was no significant difference between the growth of HT34 and HT34R. Similarly, the strain HT36 was adapted to 36 °C, as the growth of HT36R was even better than that of HT36 (Fig. 2c). Moreover, the strain HT385 was able to grow at 38.5 °C, regardless of the fact that a longer lag phase was observed (Fig. 2d). There was no significant difference between the growth of HT385R and HT385, implying the adaptation of HT385 to 38.5 °C. Collectively, these results suggest that ALE combined with mutagenesis is an efficient way to breed thermotolerant Y. lipolytica.
Fig. 2
Growth curves of Y. lipolytica CA20, Y. lipolytica HT34, Y. lipolytica HT36, and Y. lipolytica HT385 under 30 (a), 34 (b), 36 (c), and 38.5 °C (d). HT34, HT36, and HT385 were grown at 30 °C for 30 days, after which the strains were returned to grow at 34, 36, and 38.5 °C, respectively, to produce the strains HT34R, HT36R, and HT385R. Data obtained from three biological replicates were shown as the mean ± standard deviation. Different colors of “*” indicating significant differences are in accordance with the respective strain (*p < 0.05; ***p < 0.001). The control group in a, b, c, and d was set as CA20, HT34, HT36, and HT385, respectively
Genome changes of Y. lipolytica during thermal adaptive evolutionAfter obtaining the thermotolerant Y. lipolytica, we next sought to figure out how cells respond to thermal stress at a genomic level. Hence, the genomes of Y. lipolytica HT34, Y. lipolytica HT36, and Y. lipolytica HT385 were sequenced and compared with Y. lipolytica CA20 to find genome variants. The results showed that SNP, Indel, and SV were both found in the thermotolerant strains (Fig. 3a–c). The number of SNP and Indel in HT34, HT36, and HT385 was 302/358, 295/368, and 261/394, respectively (Fig. 3d). The SNP and Indel caused the nonsynonymous mutation of genes in HT34, HT36, and HT385, with a number of 10, 13, and 12, respectively (Fig. 3e). Ten mutant genes were both identified in HT34 and HT36, while only one mutant gene (A005324) was shared by HT34, HT36, and HT385 (Table 1). Moreover, mutations resulted in the deletion of 2, 3, and 7 genes in HT34, HT36, and HT385, respectively, either by the gain of the stop codon (stopgain) or frameshift. Additionally, abundant SV was found in the chromosome of HT385 compared with that of CA20, especially for the intra-chromosomal translocation (ITX) loci (Fig. 3f).
Fig. 3
Genome changes of thermotolerant strains during adaptive evolution. An overview map of the SNP, Indel, and SV sites on the chromosomes of HT34 (a), HT36 (b), and HT385 (c). The number of SNP and Indel (d), mutated genes (e), and SV (f). SNP, single-nucleotide polymorphisms; Indel, insertions and deletions; SV, structure variations; CTX, inter-chromosomal translocation; DEL, deletion; INS, insertion; INV, inversion; ITX, intra-chromosomal translocation
Table 1 Mutated genes of HT34, HT36, and HT385 during thermal adaptive evolutionFunctional annotation of the mutant genes showed that abundant genes in HT34 or HT36 were involved in transcription regulation (e.g., A003076, A004232, A001169, and A005977) and translation regulation (A002823). The mutant genes in HT385 mainly participated in signal transduction (A001479, A003939, and A005494), translational initiation and posttranslational modification (A004754, A005130, and A005261), and ATP transport (A003822). In addition, genes related to DNA repair (A001179 and A003318) and cell cycle (A001929) were found to be mutated during adaptive evolution. Collectively, these results suggest that mutation of genes involved in transcription and translation regulation is one important way for cells to adapt to the increased thermal stress.
Global gene expression of the thermotolerant Y. lipolyticaKnowing that genes involved in expression regulation were mutated during thermal adaptive evolution, we sought to deepen our understanding of the thermotolerance mechanism of these strains at a transcriptional level. To achieve this goal, the transcriptomes of Y. lipolytica CA20 (named as YLX3), Y. lipolytica HT36 (named as YLX6), and Y. lipolytica HT385 (named as YLX8) were analyzed and compared. Totally, 923 and 1162 genes were identified to be differentially expressed in YLX6 and YLX8 compared with those in YLX3, of which 653 and 888 genes were found to be upregulated, respectively (Supplemental Fig. S3). Moreover, 557 genes were differentially expressed in YLX8 compared with those in YLX6, wherein 372 genes were upregulated. The number of common DEGs found among groups YLX6 vs. YLX3, YLX8 vs. YLX3, and YLX8 vs. YLX6 was 116 (Fig. 4a and Supplemental Table S3).
Fig. 4
Functional enrichment analysis of the DEGs. a Venn diagram among groups YLX6 vs. YLX3, YLX8 vs. YLX3, and YLX8 vs. YLX6. GO enrichment analysis of the DEGs in YLX6 (b) and YLX8 (c) compared with that in YLX3. KEGG enrichment analysis of the DEGs in YLX6 (d) and YLX8 (e) compared with that in YLX3
The genes encoding hypothetical proteins with unknown functions were not discussed in this study. GO enrichment analysis showed that most of the upregulated genes in YLX6 and YLX8 encode proteins that are located at the cell periphery, extracellular region, intrinsic component membrane, and TCA complex (Fig. 4b, c). These proteins predominantly possess oxidoreductase activity, catalytic activity, and transmembrane transporter activity, functioning in bioprocesses like the oxidation–reduction process, cellular response to stimulus, and small-molecule catabolic process. The downregulated genes in YLX6 and YLX8 mainly code for proteins that are located at the cell periphery and plasma membrane, possessing oxidoreductase activity and transmembrane transporter activity, and functioning in transmembrane transport and the oxidation–reduction process (Fig. 4b, c). KEGG analysis showed that most of the DEGs found in YLX6 and YLX8 were distributed in pathways including carbohydrate metabolism, amino acid metabolism, replication and repair, lipid metabolism, signal transduction, cell growth and death, and transport and catabolism (Fig. 4d, e). We next conducted a detailed study of the transcriptional changes of these important pathways and within the functional groups revealed by the enrichment analysis.
Cell envelope structuresMembrane channels like the major facilitator superfamily (MFS) transporter and porin are critical for nutrient uptake. We found that 14 DEGs were related to the transporters (Fig. 5 and Supplemental Table S4), of which 10 genes coding for MFS-type transporter (A000353, A003235, A004474, A004837, and A005898), general substrate transporter (A000729), plasma membrane low glucose sensor (A000826), amino acid permease (A000501), monocarboxylate/proton symporter (A001982), and YALIA101S01e17150g1_1 (A000973) were upregulated in YLX8 or YLX6 compared with that in YLX3. These changes suggested that cells increased nutrient uptake to defend against thermal stress. The downregulated transporters were correlated with the transport of irons (A000881, A005124, and A005659) and water (A002359).
Fig. 5
Outline of the DEGs in YLX6 and YLX8 compared with those in YLX3. DEGs were shown to be involved in biological processes including signal pathway, lipid metabolism, fatty acid degradation, DNA replication and repair, transporter system, glycolysis, tricarboxylic acid cycle (TCA), propanoate metabolism, amino acid metabolism, oxidative stress response, and ubiquitin-mediated proteolysis. PKA, cAMP-protein kinase A; RAP1, RAS-related protein Rap-1A; RafA, A-Raf proto-oncogene serine/threonine-protein kinase; RafB, B-Raf proto-oncogene serine/threonine-protein kinase; Raf1, RAF proto-oncogene serine/threonine-protein kinase; MEK1/MEK2, mitogen-activated protein kinase 1/2; NR1C1, peroxisome proliferator-activated receptor alpha; gldA encodes glycerol dehydrogenase; DGHS, diacylglycerylhomoserine; DGTS, diacylglyceryl-N,N,N-trimethylhomoserine; galC encodes galactosylceramidase; GABA, γ-aminobutyric acid; P, phosphate. An arrow with a dashed line indicates that the corresponding process includes more than one step. The gene names with red and green text indicate upregulation and downregulation, respectively. Detailed Log2FC (fold change) values, together with the gene annotations, were listed in the Supplemental Table S4
In addition to the transporters, thermal stress also caused the differential expression of genes involved in lipid metabolism (Fig. 5). Notably, the DEGs involved in steroid biosynthesis (A000926, A000927, A000928, A001412, and A003411), glycerolipid and glycerophospholipid metabolism (A002374, A003797, and A004202), and ceramide synthesis (A003183 and A004782) were upregulated in YLX6 and YLX8 compared with that in YLX3, suggesting that alteration of the membrane lipid component was one important way for cells to grow under thermal stress.
Glycolysis and tricarboxylic acid cycleTen DEGs were identified to be related to glycolysis and the TCA cycle (Fig. 5). Interestingly, the expression of genes coding for phosphoglucose isomerase (A005351), fructose-1, 6-bisphosphate (FBP) aldolases (A004415), tetrameric phosphoglycerate mutase (A001768), enolase (A005710), pyruvate kinase (A005412), pyruvate dehydrogenase beta subunit (A004455), and TPP-dependent 2-oxoacid decarboxylase (A005245) decreased in YLX6 compared with that in YLX3 (Supplemental Table S4), suggesting that the glycolysis was inhibited upon thermal stress. Moreover, the expression of genes related to the TCA cycle, including A002777 and A003610 coding for malate dehydrogenase and isocitrate dehydrogenase, respectively, was also downregulated in YLX6 compared with that in YLX3, indicating a suppressed TCA cycle of cells upon thermal stress.
Amino acid metabolism, fatty acid degradation, and propanoate metabolismThe inhibited glycolysis might result in the reduced synthesis of acetyl-CoA, thereby influencing the TCA cycle. Cells might metabolize other molecules to supply acetyl-CoA and maintain the running of the TCA cycle. We found that 16 DEGs were involved in the amino acid metabolism and propanoate metabolism, of which transcriptional upregulation mainly occurred in the genes related to the metabolism of leucine, valine, and isoleucine (A002420, A002522, A003071, A003895, A003896, A004839, A005184, A005940, and A006115), the metabolism of glycine, threonine, and serine (A001367 and A006263), and the metabolism of propanoate (A005184 and A005474) in YLX6 and YLX8 compared with that in YLX3 (Fig. 5 and Supplemental Table S4). Besides, four genes involved in fatty acid degradation (A002282, A003667, A004479, and A004709) were shown to be upregulated upon thermal stress. In addition, the expression of genes involved in glutamate metabolism was downregulated in YLX6 and YLX8 compared with that in YLX3, including A001092, A002654, A005380, and A005754, suggesting an accumulation of glutamate in the cells. These results suggest that cells metabolize leucine, valine, isoleucine, fatty acid, and propanoate to supply acetyl-CoA and propanoyl for the normal running of the TCA cycle while accumulating glutamate to defend against thermal stress.
DNA replication, repair, and recombinationMaintenance of genome stability is important for cells to survive in stressful environments. Notably, we found that among the 12 DEGs related to DNA replication, 10 genes (A000180, A000201, A000385, A001165, A001477, A002480, A004005, A004732, A005285, and A006185) mainly coding for nucleoside triphosphate hydrolase, DNA polymerase, replication factor, and DNA primase were upregulated in YLX6 and YLX8 compared with that in YLX3 (Fig. 5 and Supplemental Table S4). Some of these genes were also involved in DNA repair. Additionally, transcriptional upregulation was observed for the genes included in nucleotide excision repair (A004466), mismatch repair (A004466 and A006112), base excision repair (A002035 and A004995), and homologous recombination (A001013 and A005661). These results suggest that cells increase genome stability to survive under thermal stress.
Oxidative stress response and signaling pathwayTo withstand the oxidative stress caused by high temperature, cells increased the expression of genes involved in peroxisome. Among the 17 DEGs, the expression of 14 genes coding for hydrolase protein (A000184, A004170, and A005374), peroxisome biogenesis factor (A000525 and A002350), catalase (A006279), choline/carnitine o-acyltransferase (A005901), deoxycytidine monophosphate (dCMP) deaminase (A002252), peroxisomal 2, 4-dienoyl-CoA reductase (A003958), FAD/FMN-dependent oxidoreductase (A002638 and A006263), peroxisome assembly protein 12 (A003191), acyl-CoA oxidase (A003667), and peroxisomal 3-oxoacyl-CoA thiolase (A004119) was upregulated, while 3 genes coding for catalase (A003253 and A003272) and redoxin (A002165) were downregulated (Fig. 5 and Supplemental Table S4). Moreover, the expression of genes coding for WD40-repeat-containing protein (A000608) and ubiquitin-conjugating enzyme (A001803 and A002553) was upregulated, indicating that oxidative stress induced protein aggregation and misfolding. The RAS-controlled signal pathway (cAMP-protein kinase A (PKA) pathway) plays a positive role in the control of dimorphic transition in yeast (Liang et al. 2017). We found that the expression of two genes coding for RAS-related proteins (A001014 and A005979) was increased in YLX6 and YLX8 compared with that in YLX3, suggesting an involvement of the RAS-cAMP-PKA signal pathway in the thermal stress response.
Erythritol metabolismErythritol could be synthesized from glucose through the pentose phosphate pathway (Fig. 6a). Here, we found that 11 DEGs were involved in erythritol metabolism, among which A004280 coding for glucose-6-phosphate dehydrogenase (ZWF1), A001342 coding for phosphogluconate dehydrogenase (GND1), A003663 coding for transketolase (TKL1), A005663 coding for transaldolase (TAL1), A005140, A001913, and A005791 coding for erythrose reductase (ER10, ER25, and ER27) were responsible for erythritol synthesis, while A002376 coding for erythritol dehydrogenase (EYD1), A002374 coding for erythrulose kinase (EYK1), A002373 coding for erythrulose-1-phosphate isomerase (EYI1), and A002375 coding for erythrulose-4-phosphate isomerase (EYI2) were involved in erythritol degradation (Fig. 6a and Supplemental Table S4). Notably, almost all of the genes involved in erythritol synthesis were downregulated in YLX6 and YLX8 compared with those in YLX3, while the genes related to erythritol degradation were upregulated, suggesting a decreased accumulation of erythritol in HT36 and HT385 (Fig. 6b, c). Expectedly, the titer and yield of erythritol were significantly decreased in HT36 and HT385 compared with those of CA20 (Fig. 6d, e).
Fig. 6
DEGs involved in erythritol metabolism. a Erythritol metabolism pathway. b Changes in the expression level of genes in YLX6 compared with that of YLX3. c Changes in the expression level of genes in YLX8 compared with that of YLX3. d The production of erythritol by Y. lipolytica CA20, Y. lipolytica HT36, and Y. lipolytica HT385 at different temperatures. e The erythritol yield. GK, glucokinase; HK, hexokinase; ZWF1, glucose-6-phosphate dehydrogenase; PGLS, 6-phosphogluconolactonase; GND1, phosphogluconate dehydrogenase; RPE1, ribulose phosphate 3-epimerase; RPI, ribulose-5-phosphate isomerase; TKL1, transketolase; TAL1, transaldolase; E4PK, erythrose-4-phosphate kinase; ER, erythrose reductase; EYD1, erythritol dehydrogenase; EYK1, erythrulose kinase; EYI1, erythrulose-1-phosphate isomerase; EYI2, erythrulose-4-phosphate isomerase; P, phosphate. The green and red text represent downregulation and upregulation, respectively. Data obtained from three biological replicates were shown as the mean ± standard deviation. “*” indicates that there is a significant difference between the two studied groups (***p < 0.001), in which CA20 is set as the control group
Alteration of SOD and CAT activity, the content of intracellular FFA, AA, and ATP in Y. lipolytica HT385SOD and CAT are essential for cells to defend against oxidative stress. Interestingly, at a transcriptional level, the genes coding for SOD were not found to be differentially expressed in YLX8 compared with that in YLX3, while two of the three DEGs coding for catalase were downregulated (Supplemental Table S4). We next detected the SOD and CAT activity of Y. lipolytica HT385 and Y. lipolytica CA20 throughout the growth phase. The results showed that the SOD activity of HT385 was significantly higher than that of CA20 when grown for 24 h, after which the SOD activity of HT385 and CA20 decreased as cells grew, and there was no significant difference observed between the two groups at 48 and 72 h (Fig. 7a). A similar phenomenon was observed for the changes of CAT activity in HT385 and CA20 (Fig. 7b). These findings overall supported the results obtained from the transcriptome analysis.
Fig. 7
Alteration of the SOD activity (a), CAT activity (b), FFA content (c), AA content (d), ATP content (e), and CFU (f) of Y. lipolytica HT385 and Y. lipolytica CA20 throughout the whole growth phase. Data obtained from three biological replicates were shown as the mean ± standard deviation. “*” indicates that there is a significant difference between the two studied groups (**p < 0.01; ***p < 0.001). SOD, superoxide dismutase; CAT, catalase; FFA, free fatty acid; AA, amino acid; CFU, colony-forming units
For the changes of FFA content, we found that the FFA content of HT385 and CA20 decreased gradually as cells grew, with the FFA content of HT385 being significantly higher than that of CA20 within 48 h while lower than that of CA20 at 72 h (Fig. 7c). Similarly, the AA content of HT385 decreased as cells grew and was significantly higher than that in CA20 within 48 h (Fig. 7d). These results appeared to support the conclusion deduced from the transcriptome analysis that cells metabolized FFA and AA to increase the synthesis of acetyl-CoA to defend against thermal stress. Additionally, the ATP content of HT385 was significantly higher than that in CA20 throughout the growth phase (Fig. 7e), emphasizing the importance of ATP in the thermal stress response. CFU detection showed that the cell number of HT385 was lower than that of CA20 at 24 h (Fig. 7f), which was consistent with the growth curve of HT385 at 38.5 °C, wherein a lag phase was observed within 24 h (Fig. 2d).
Influence of glycerol, soybean oil, trehalose, AA, thiamine, and ATP on the growth of Y. lipolytica HT385After knowing that FFA and AA metabolism contributed to the thermotolerance of Y. lipolytica HT385, we sought to figure out whether the exogenous addition of these substances could enhance the growth of cells. Meanwhile, the effect of glycerol, thiamine, trehalose, and ATP on the growth of HT385 was determined as well. The results showed that the addition of glycerol significantly suppressed the growth of HT385 compared with the control group (WA), and the inhibitory effect was enhanced as the glycerol concentration increased (Fig. 8a). Soybean oil contains abundant fatty acids. It was found that addition of soybean oil with a final concentration of 5 or 10 g/L produced little influence on the cell growth within 48 h, whereas soybean oil of 20 g/L significantly promoted the cell growth compared with that of WA and also shortened the lag phase within 24 h (Fig. 8b). The addition of trehalose with a final concentration of either 0.5, 1, or 2 g/L had little effect on the growth of cells (Fig. 8c). Similar results were observed for the addition of glutamate, wherein 1, 10, and 20 μM glutamate were not able to promote cell growth (Fig. 8d). In addition, it was worth noting that the addition of either leucine, isoleucine, or valine could significantly promote cell growth compared with that of WA in most cases (Fig. 8e–g). Moreover, the addition of leucine, isoleucine, and valine could increase the maximum cell density. Additionally, we found that the addition of thiamine or ATP with an interval of 12 h was not able to facilitate cell growth (Fig. 8h, i). Collectively, these results further support that FFA, leucine, isoleucine, and valine are critical for cells to grow under thermal stress.
Fig. 8
The influence of glycerol (a), soybean oil (b), trehalose (c), glutamate (d), leucine (e), isoleucine (f), valine (g), thiamine (h), and ATP (i) on the growth of Y. lipolytica HT385. Data obtained from three biological replicates were shown as the mean ± standard deviation. Different colors of “*” indicating significant differences are in accordance with the respective strain to the control group (*p < 0.05; **p < 0.01; ***p < 0.001). WA, without addition; arrows indicate that the thiamine or ATP is added at the corresponding time points
Effect of the overexpression of upregulated genes on the growth and erythritol production of Y. lipolytica CA20Transcriptome analysis showed that abundant genes coding for hypothetical proteins with unknown functions were upregulated in thermotolerant strains, indicating the important roles of these genes in the thermal stress response. To test this hypothesis, eighteen representative genes with a relatively higher expression level were selected and overexpressed in Y. lipolytica CA20 (Supplemental Table S5). Among the eighteen genes, three were related to ceramide synthesis (A003183) and the peroxisome (A000121 and A006279), while other genes mainly encode hypothetical proteins. We found that overexpression of either A000184, A000800, A001678, A002375, A002808, A004625, A004733, or A006279 had little effect on the growth of CA20 at 30 °C, as no significant difference in cell growth was observed between the CA20∆ura3 harboring a target gene and the vector pDCXRA throughout the growth phase (Fig. 9a, b). On the contrary, individual overexpression of the genes A002175, A003183, A003902, A004055, A004467, A004535, A005690, A005844, and A006220 significantly promoted cell growth within 24 h compared with that observed in CA20∆ura3::pDCXRA (Fig. 9c, d). In addition, overexpression of A000121 improved cell growth after 48 h.
Fig. 9
Growth curves of CA20∆ura3 with an overexpression of the upregulated genes at 30 °C (a–d) and 34 °C (e, f). The YNBD medium was used in the growth assay. Data obtained from three biological replicates were shown as the mean ± standard deviation. Different colors of “*” indicating significant differences are in accordance with the respective strain to the control group (*p < 0.05; **p < 0.01; ***p < 0.001). The comparison was conducted between the CA20∆ura3 harboring a target gene and the vector pDCXRA
Further, we investigated whether overexpression of the selected genes could enable cells to grow at a temperature higher than 30 °C. It was found that overexpression of A000121, A002175, A002375, A003183, A003902, A004055, A004467, A004535, A005690, A005844, and A006220 individually enabled the cells to grow at 34 °C, while CA20∆ura3 harboring the vector pDCXRA was not able to grow (Fig. 9e, f). Notably, CA20∆ura3 with an overexpression of either A000121, A003183, or A005690 grew better than other strains at 34 °C, suggesting that these genes would be good candidates for the construction of thermotolerant strains by genetic engineering. Further, the growth of these strains at 35, 36, and 38.5 °C was tested. It was found that cells with an overexpression of these genes were not able to grow at 35 °C (Supplemental Fig. S4) or higher temperatures (data not shown).
Effect of overexpression of the upregulated genes on erythritol production at 30 and 34 °C was further investigated. The results showed that individual overexpression of the most genes had little effect on the erythritol production by CA20∆ura3 at 30 °C, as no significant difference in the erythritol titer was observed between the CA20∆ura3 harboring a target gene and the vector pDCXRA (Fig. 10). Overexpression of A002375 significantly decreased the erythritol titer, while individual overexpression of A003183 and A005690 significantly promoted the synthesis of erythritol by CA20∆ura3. At 34 °C, the production of erythritol was inhibited in CA20∆ura3::pDCXRA compared with that obtained at 30 °C. Similarly, overexpression of A002375 also significantly decreased the erythritol titer. Nevertheless, the erythritol production of CA20∆ura3 with an individual overexpression of A000121, A003183, A005844, and A006220 was significantly increased compared with that of CA20∆ura3::pDCXRA. The erythritol titers of these strains obtained at 30 °C and 34 °C were similar, as no significant difference was observed. Moreover, the erythritol titer of CA20∆ura3::A005690 was increased and even significantly higher than that obtained at 30 °C (Fig. 10). Collectively, these results suggest that overexpression of some upregulated genes could not only endow cells with thermotolerance but also preserve erythritol production.
Fig. 10
Effect of overexpression of the upregulated genes on erythritol production by CA20∆ura3 at 30 °C and 34 °C. The erythritol production was carried out using YNBD medium containing 200 g/L glucose. Data obtained from three biological replicates were shown as the mean ± standard deviation. “*” indicates a significant difference (*p < 0.05; **p < 0.01; ***p < 0.001)
留言 (0)