
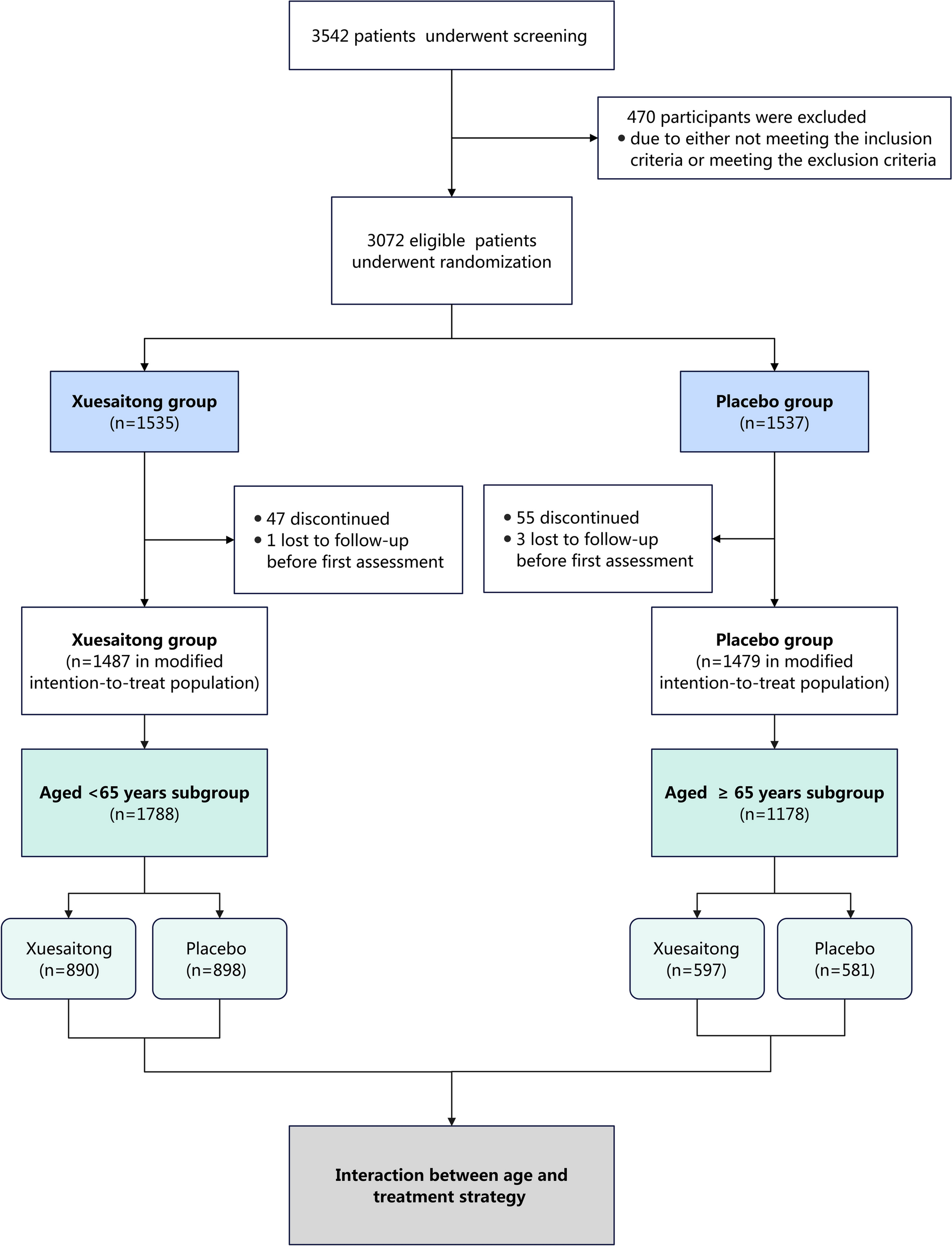
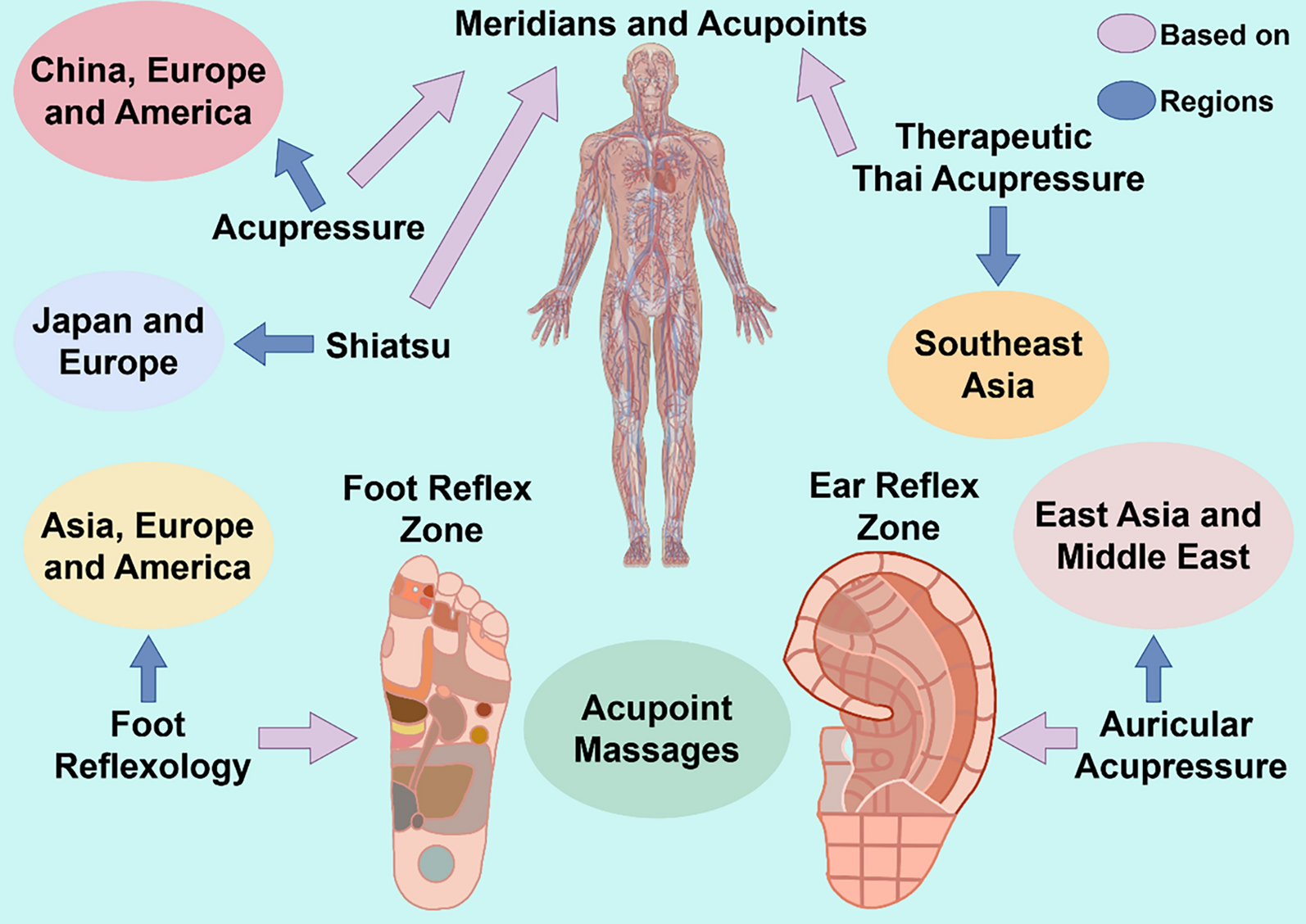
Remember me








The methodology of GBP for the prevention of drunkenness and ALD is depicted in Fig. 1A, which outlines the experimental protocol. As shown in Fig. 1B, 92% of the vehicle group exhibited drunkenness, whereas treatment with metadoxine (300 mg/kg) resulted in a 69% reduction in the incidence of drunkenness. Similarly, compared with those in the vehicle group, the proportions of mice in the low-dose (100 mg/kg) and high-dose (400 mg/kg) GBP groups decreased to 67% and 55%, respectively, indicating dose-dependent decreases in drunkenness rates of 22%, 25%, and 37%, respectively, compared to those in the vehicle group. As shown in Fig. 1C, the latency to drunkenness was 14.53 ± 11.73 min in the vehicle group and 44.25 ± 30.03 min in the positive control group. The low- and high-dose GBP groups demonstrated an even more pronounced extension of latency periods, 41.63 ± 29.43 and 128.2 ± 65.08 min, respectively. This represents latency periods increased by a factor of approximately 2.86 and 8.83 compared to those of the vehicle group. Furthermore, the duration of drunkenness, which was measured from the loss of the righting reflex, was noted at 315.73 ± 57.00 min in the vehicle group. In contrast, the positive control group experienced a significant reduction in duration to 272.93 ± 68.51 min, constituting a 14% decrease. The low- and high-dose GBP groups had durations of 290.03 ± 87.37 min and 163.98 ± 74.53 min, respectively, marking reductions of 8% and 48%, respectively (Fig. 1D). Similarly, compared with those in the vehicle group, the hepatic ALDH enzyme levels in the positive control group were by 14%, while those in the low- and high-dose GBP groups were by 11% and 21%, respectively (Fig. 1E). GBP treatment at high doses (400 mg/kg) led to noteworthy outcomes, as depicted in Fig. 1B, C, D, E. There was a significant decrease in the percentage of drunkenness, an extension in the latency period for drunkenness, a reduction in the duration of drunkenness, and an increase in hepatic ALDH enzyme levels in GBP treatment (400 mg/kg) group compared to the vehicle group (P < 0.05). Collectively, these findings affirm the potency of GBP in curtailing alcohol-induced impairment, underscoring its preventive efficacy against drunkenness and ALD.
Fig. 1
GBP prevents drunkenness in mice. A Study design for assessing the impact of GBP in alcohol treated mice. B Reduced incidence of drunkenness in GBP treatment groups. C Increased latency to drunkenness with GBP administration in mice. D Duration of drunkenness decreased upon GBP treatment. E Increased ALDH enzyme levels following GBP exposure. The data are expressed as the mean ± SD (n = 10–13). Compared with the control group, #P < 0.05; compared with the vehicle, *P < 0.05
Effects of GBP on ALD in micePathological changes in the liver are one of the primary organs reflecting the safety of a drug. To determine the protective effect of GBP on acute alcoholic liver injury, the following pathological analysis was conducted. As shown in Fig. 2A, the livers of mice in the blank group appeared healthy and normal with neatly arranged hepatocytes, no vacuolar lesions, no inflammatory cell infiltration, and no necrosis. In contrast, compared with those in the control group, the vehicle group exhibited disorganized hepatocyte arrangement, significant cell swelling, blurred cell boundaries, and an increased number of necrotic hepatocytes and fatty vacuoles. The liver scoring results are shown in Fig. 2B. Compared with those in the vehicle group, the positive control group and GBP group exhibited significant improvements in terms of well-preserved liver cell structure, clear cell boundaries, and almost no vacuolar or necrotic changes (P < 0.05). The mouse body weight data, as shown in Fig. 2C, indicated that the body weight steadily increased during the experiment. When acute liver injury occurs, the liver exhibits noticeable swelling. As shown in Fig. 2D, compared with that in the control group, the liver index of the vehicle group was significantly increased (P < 0.05), and GBP slightly reversed these changes but not significantly (P > 0.05) compared to those in the control group. Collectively, our results demonstrated that GBP effectively improved alcoholic liver injury in mice.
Fig. 2
GBP ameliorates alcohol-induced liver injury in mice. A Representative liver tissue morphology and histopathological sections. B Quantification of liver tissue pathological score, C body weight, and D liver index. The serum levels of ALT E, AST F, AKP G, TG H, TC I, and the activity of SOD in liver tissues J were evaluated. The data were expressed as the mean ± SD (n = 10–13). Compared with the control group, #P < 0.05; compared with the vehicle, *P < 0.05
Effects of GBP on the serum and liver biochemical marker levels in ALD miceTo ascertain the effect of GBP on serum biochemical markers in a mouse model of ALD, we evaluated biochemical indicators such as AST, ALT, AKP, TG, TC, and hepatic SOD. The serum ALT, AST and AKP levels, which are principal biochemical markers reflecting liver function, were significantly greater in the vehicle group than in the control group (P < 0.05), confirming the successful establishment of the ALD model in mice. In contrast, GBP group exhibited significant reductions in ALT, AST and AKP levels (Fig. 2E, F and G, P < 0.05). TG and TC, key indicators of lipid changes in the liver, were significantly elevated in the vehicle group compared to those in the control group (P < 0.05), indicating that alcohol disrupts lipid metabolism, leading to lipid accumulation. A high dose of GBP significantly decreased the levels of TG and TC (Fig. 2H, I, P < 0.05), suggesting that GBP may accelerate triglyceride metabolism to reduce fat accumulation. SOD, an antioxidative biomarker, reflects the capacity of tissue to resist oxidative damage. As illustrated in Fig. 2J, the antioxidative capacity was significantly enhanced in the high-dose GBP group than in the vehicle group (P < 0.05). Our findings indicate that GBP possesses a robust hepatoprotective effect on ALD mice.
The gut microbiota landscape and its correlation with liver biochemistryTo examine changes in the gut microbiome composition in mice after GBP treatment, we sequenced the V3-V4 region of the 16S rRNA gene from fecal samples. Venn diagrams were constructed to analyze the commonalities in microbial richness across the groups. As depicted in Fig. 3A, there was an overlap of 702 OTUs among the three groups: 1045 OTUs in the control group, 1018 OTUs in the vehicle group, and 1011 OTUs in the GBP group. Beta diversity was analyzed using PCoA. As shown in Fig. 3B, the gut microbial communities of the mice that consumed ethanol were different from those of the control group or the vehicle group. Concurrently, PCoA revealed a distinct separation pattern of gut microbial communities in mice treated with GBP. At the phylum level, Bacteroidetes, Firmicutes, Proteobacteria, and Verrucomicrobia were the predominant phyla (Fig. 3C). Compared to those in the control group, the vehicle group exhibited increased relative abundances of Proteobacteria and Patescibacteria, whereas the relative abundances of Firmicutes, Epsilonbacteraeota, Actinobacteria, Tenericutes, and Cyanobacteria were lower. However, GBP reversed this trend. At the genus level, the relative abundances of Bacteroides, Enterobacter, and Escherichia-Shigella in the vehicle group exhibited an increase compared to those in the control group, while the relative abundances of Desulfovibrio and Lachnospiraceae NK4A136_group showed a reduction. However, the administration of GBP reversed these changes (Fig. 3D). We identified 25 taxonomic groups at the gene level within the gut microbiota that were significantly influenced by GBP. Among these 25 microbial taxa, Escherichia-Shigella was found to be significantly enriched in the vehicle group and was positively correlated with ALT, AKP, TG and TC but negatively correlated with ALDH. On the other hand, the abundance of Lachnospiraceae NK4A136 decreased significantly in the vehicle group, with negative correlations with ALT, AKP and TG and a positive correlation with SOD. However, supplementation with GBP reversed these changes (Fig. 3E). These data collectively suggest that GBP modulates the gut microbiota, which may be correlated with improvements in hepatic function.
Fig. 3
Modulation of the gut microbiota and correlation with liver biochemistry in response to GBP treatment. A Venn diagram showing shared and unique OTUs among the control, Vehi, and GBP groups. B PCoA plot visualizing beta diversity, with greater distances indicating more distinct microbiota compositions. C Bar chart detailing the relative abundance of microbial taxa at the phylum level among the different groups. D Bar chart displaying the relative abundance at the genus level. E Heatmap of genus-level abundance and correlation with key hepatic biochemical parameters. The sample size of each group was 8. The blue squares indicate a significant positive correlation, and the red squares indicate a significant negative correlation (P < 0.05)
To elucidate the key active microbial taxa in our study, we employed LEfSe analysis to identify biomarkers at various taxonomic levels (with LDA scores [− log10] > 3.5). Figure 4A reveals the identification of 24, 7, and 17 distinct biomarkers in the control, vehicle, and GBP groups, respectively. Figure 4B illustrates the distribution of different species across levels, with the size of each circle indicating species richness. At the phylum level, biomarkers identified in the control group included Verrucomicrobia, Epsilonbacteraeota, and Deferribacteres, whereas Proteobacteria emerged as a significant biomarker in the vehicle group. Notably, the GBP group exhibited no discernible phylum-level differences. At the genus level, the control group was characterized by the presence of biomarkers such as Akkermansia, Helicobacter, Mucispirillum, Parasutterella, and Enterorhabdus. In contrast, the biomarkers identified in the vehicle group were limited to Escherichia_Shigella and Enterobacter, while the biomarkers identified in GBP group included Lachnospiraceae NK4A136_group, Parabacteroides, Turicibacter, Romboutsia, and Alistipes. Statistical analysis of the intergroup differences revealed that alcohol consumption significantly decrased the community richness of the Lachnospiraceae NK4A136 group and increased the abundance of Escherichia-Shigella. In contrast, GBP administration markedly promoted intestinal microbial homeostasis, as demonstrated in Fig. 4C.
Fig. 4
Comparative analysis of microbial communities among the groups. A The histogram displays LDA scores for features with differential abundance among the various groups. The scores indicate the magnitude of the effect size corresponding to each feature in distinguishing among groups. Taxa listed on the y-axis with LDA scores exceeding 3.5 are highlighted as potential biomarkers for their respective groups. B A cladogram illustrating the phylogenetic distribution of the microbiota determined by LEfSe analysis, with circle layers representing taxonomic classifications from kingdom to genus. Nodes are color-coded based on the group with the most pronounced association with each taxon. C A bar graph shows the statistical evaluation of genus-level differences among the control, vehicle, and GBP groups. The sample size in both groups was 8. Compared with the control group, #P < 0.05; compared with the vehicle, *P < 0.05
Identifying the ingredients of GBP in mouse serum via mass spectrometryThe UNIFI 1.8.2 (Waters Corp., Milford, USA) platform was utilized to process and analyze the MS data, involving database matching, mass accuracy, isotope patterns, and fragmentation pathways (sourced from the literature and public data). The total ion chromatogram of GBP is shown in Fig. 5. In total, 7 ingredients (3-keto tauroursodeoxycholic acid, 7-ketodeoxycholic acid, cholic acid, taurocholic acid, taurochenodeoxycholic acid, ursodeoxycholic acid, and chenodeoxycholic acid) were identified, and their mass spectrometry details are provided in Additional file 1: Table S1.
Fig. 5
UPLC-Q/TOF–MS chromatographic analysis of GBP ingredients and metabolites. The chromatograms display the retention times (min) of each peak on the x-axis, with the y-axis indicating ion counts that signify the relative abundance of each ingredient or metabolite. The upper chromatogram shows data in positive ion mode, whereas the lower chromatogram shows results in negative ion mode
Multivariate statistical analysis and metabolite identificationThe OPLS-DA score plots distinctly segregated the control, vehicle, and GBP treated groups (Fig. 6A). We identified 39 potential metabolic features significantly influenced by GBP (P < 0.05, FC > 2, and VIP > 1). Four crucial metabolic pathways, namely glycerophospholipid metabolism, alpha-linolenic acid metabolism, retinol metabolism, and caffeine metabolism, were identified based on − log (P) > 1 and an impact value > 0.02 (Fig. 6B). Among these 39 metabolites (Additional file 2: Table S2), phospholipids (PCs, PEs, PIs, LysoPCs, and LysoPEs) were significantly enhanced in the vehicle group and positively correlated with TC and TG. Moreover, fatty acids (behenic acid, erucic acid, and docosatrienoic acid) were significantly decreased in response to GBP treatment and were positively correlated with ALT, TG, and TC. Additionally, two bile acids, alpha-muricholic acid (α-MCA) and hyodeoxycholic acid (HDCA), were positively correlated with the levels of ALT and AKP. In contrast, the concentrations of SOD and ALDH were negatively associated with these compounds. These parameters increased in the Vehicle group, but GBP treatment significantly reversed this trend. Furthermore, Taurocholic acid (TCA) was negatively correlated with the contents of ALT, AKP, TG, TC, which decreased in the vehicle mice, respectively (Fig. 6C). Taken together, these results suggest that GBP intervention alters the serum metabolite levels and is correlated with the regulation of biomarkers of hepatic function.
Fig. 6
Metabolic profiling and correlation with hepatic biochemical indices in response to treatment. A OPLS-DA score plots illustrating the metabolic differences between the control (green), vehicle (blue), and GBP (orange) groups in negative and positive ionization modes. B A volcano plot showing metabolites exhibiting significant intergroup differences, with the size reflecting the magnitude of change and the color denoting P-value indicating statistical significance. C A heatmap representing z-score normalized metabolite intensities in the different groups and their correlation with hepatic biochemistry parameters (ALT, AST, AKP, TG, and TC). The sample size of each group was 8. The blue squares indicate a significant positive correlation, and the red squares indicate a significant negative correlation (P < 0.05)
Correlations between various metabolites and the gut microbiotaThe potential correlation between the gut microbiota and metabolites was investigated in depth by calculating Spearman’s rank correlation coefficient. A correlation analysis between the 39 differentially abundant metabolites and the 29 distinct microbiota communities at the genus level was performed. This analysis revealed significant correlations (P < 0.05) between bacteria and different metabolites, which were visually represented in a heatmap (Fig. 7). Rikenella, Mucispirillum, and Parasutterella exhibited a negative correlation with phospholipids, whereas Enterobacter, Parabacteroides, Helicobacter, and Alistipes demonstrated a positive correlation with these compounds. Lachnospiraceae, Lachnoclostridium, and Alistipes positively interact with α-MCA and HDCA, while Faecalibaculum negatively interacts with these substances. These findings indicate a correlation between alterations in the gut microbiota and specific metabolites.
Fig. 7
Heatmap visualizing the correlations between metabolites and the gut microbiota. The heatmap employs Spearman’s rank correlation coefficients to display the relationships, with color gradations indicating the correlation strength: red signifies a positive correlation, blue signifies a negative correlation, and the color saturation denotes the magnitude, with deeper hues indicating stronger correlations (P < 0.05). The sample size of each group was 8
Comments (0)