
記住我

The olivo-cerebellar system plays an important role in vertebrate sensorimotor control. Here, we investigate sensory representations in the inferior olive (IO) of larval zebrafish and their spatial organization. Using single-cell labeling of genetically identified IO neurons, we find that they can be divided into at least two distinct groups based on their spatial location, dendritic morphology, and axonal projection patterns. In the same genetically targeted population, we recorded calcium activity in response to a set of visual stimuli using two-photon imaging. We found that most IO neurons showed direction-selective and binocular responses to visual stimuli and that the functional properties were spatially organized within the IO. Light-sheet functional imaging that allowed for simultaneous activity recordings at the soma and axonal level revealed tight coupling between functional properties, soma location, and axonal projection patterns of IO neurons. Taken together, our results suggest that anatomically defined classes of IO neurons correspond to distinct functional types, and that topographic connections between IO and cerebellum contribute to organization of the cerebellum into distinct functional zones.
Significance StatementUsing the transparent larval zebrafish, we systematically recorded the responses of inferior olive (IO) neurons to visual motion stimuli that drive optomotor and optokinetic behaviors. We find that most IO neurons respond selectively to one or more such stimuli. Individual neurons are tuned to specific directions of motion and different functional types are distributed nonuniformly in the IO. Furthermore, we were able to link the functional type of the IO neurons with their location, morphology, and projection patterns. This shows how topographically organized projections from the IO play an important role in channeling behaviorally relevant information to the cerebellum in zebrafish larvae.
IntroductionThe olivo-cerebellar system plays an important role in sensorimotor control and coordination in vertebrates. The inferior olive (IO) sends climbing fiber (CF) projections to the cerebellum, where each CF makes extensive excitatory synaptic connections with a single Purkinje cell in such a way that a single presynaptic action potential elicits a characteristic postsynaptic complex spike (Eccles et al., 1966). These complex spikes play a fundamental role in modulating Purkinje cell simple spikes and thus fine-tune cerebellar output (Marr, 1969; Albus, 1971; Ito et al., 1982).
Studies in mammals have identified a population of IO neurons that receives input from direction-selective cells in the accessory optic system, a visual pathway mediating the detection of optic flow (Soodak and Simpson, 1988). This population conveys visual sensory error signals to the cerebellum in the form of retinal slip, giving rise to complex spikes that are direction selective, and out of phase with simple-spike firing rate (Ito, 1982; Stone and Lisberger, 1990). In addition to direction selectivity, electrophysiology studies have found evidence of functional organization of CF projections. For instance, CFs carrying motion signals consistent with rotation about horizontal or vertical axes project to distinct zones of the cerebellar flocculus (Schonewille et al., 2006; Pakan et al., 2011). Moreover, in pigeons, cerebellar zones responding to different types of optic flow stimuli (Wylie and Frost, 1993; Wylie et al., 1993) have been shown to receive input from different regions of the IO (Craciun et al., 2018).
Recent calcium imaging of Purkinje cells in zebrafish larvae has revealed distinct areas of the cerebellum associated with motion stimuli that drive distinct visuomotor behaviors: the optomotor response (OMR), which drives the fish to swim and turn in the direction of visual motion, and the optokinetic reflex (OKR), which drives the eyes to track the direction of the rotation and make rapid resetting saccades in the opposite direction (Matsui et al., 2014; Knogler et al., 2019). Additionally, electrophysiological recordings from larval zebrafish Purkinje cells, located in different regions of the cerebellum, showed that complex spike responses can be grouped into different visual categories, related with changes in luminance and direction-selective translational or rotational motion (Knogler et al., 2019). Since translational and rotational motion stimuli drive different visually driven motor behaviors in larval zebrafish (Neuhauss et al., 1999), these results have led to the proposed separation of the zebrafish cerebellum into different behavioral modules. According to this model, activity in the medial cerebellum is associated with swimming and turning movements during the OMR, the medial–lateral cerebellum processes changes in luminance, and the lateral cerebellum is involved in eye and body coordination during the OKR (Matsui et al., 2014; Knogler et al., 2019). Distinct functional mapping across medial–lateral cerebellar regions in larval zebrafish is further supported by anatomical mapping of cerebellar outputs (Heap et al., 2013; Takeuchi et al., 2015; Kunst et al., 2019).
Taken together, these studies suggest that spatially segregated and functionally distinct Purkinje cells can differentially modulate various aspects of sensorimotor control and coordination. However, the extent to which a topographic organization is already present in the zebrafish IO and is fed forward by topography-preserving CF projections, or whether a different principle organizes the representations in the IO, is still unknown. This is partly because most functional studies have focused on Purkinje cell complex spikes as a proxy for IO activity, due to technical challenges associated with recording activity from IO neurons in behaving animals.
In this study, we take advantage of the larval zebrafish’s small, transparent brain in combination with genetic tools to investigate the IO, to better understand its structural and functional organization at a cellular and population level. We show that IO neurons can be divided into at least two distinct anatomical types based on their spatial location, dendritic morphology and axonal projection patterns. Furthermore, most IO neurons respond to visual stimuli that drive distinct behaviors in a direction-selective manner, with functional properties spatially organized within the IO. We describe both anatomical and functional segregation of IO neurons that can be associated with different cerebellar modules, relevant for visually driven behaviors such as the OMR and OKR.
Materials and MethodsExperimental design and statistical analysisThis study was aimed to characterize morpho-anatomical and functional properties of the IO neurons in larval zebrafish in order to understand whether the topological organization described in the zebrafish cerebellum is already present in its presynaptic source, the IO. To this end, we designed the following four experiments:
Experiment 1: Morpho-anatomical characterization of the IO neurons (Fig. 1). In this experiment we used 39 zebrafish larvae to label 53 individual IO neurons and to characterize their dendritic morphology, anatomical location within the IO, and projection patterns within the cerebellum.
Experiment 2: Characterization of the IO neuronal activity in response to whole-field translational and rotational visual motion (Fig. 2). Such stimuli were chosen because they are known to induce distinct behavioral responses in larval zebrafish and are associated with activity in distinct anatomical regions in the cerebellum (Matsui et al., 2014). In this experiment, we recorded activity of 1,106 IO neurons from 12 animals.
Experiment 3: Characterization of the IO neuronal activity in response to monocular stimulation (Fig. 3). In this experiment, we aimed to understand how (if at all) the responses observed in Experiment 2 result from binocular integration of monocular inputs. To this end, we recorded activity of 518 IO neurons from six animals.
Experiment 4: Linking the morpho-anatomical and functional organization of the IO (Fig. 4). To this end, we recorded binocular responses from 28 larvae not only from the somata of the IO neurons but also from their axon terminals within the cerebellum using light-sheet microscopy. This approach allowed us to directly link the morpho-anatomical and functional properties of the IO neurons with the functionally compartmentalized organization of the zebrafish cerebellum.
 Figure 1.
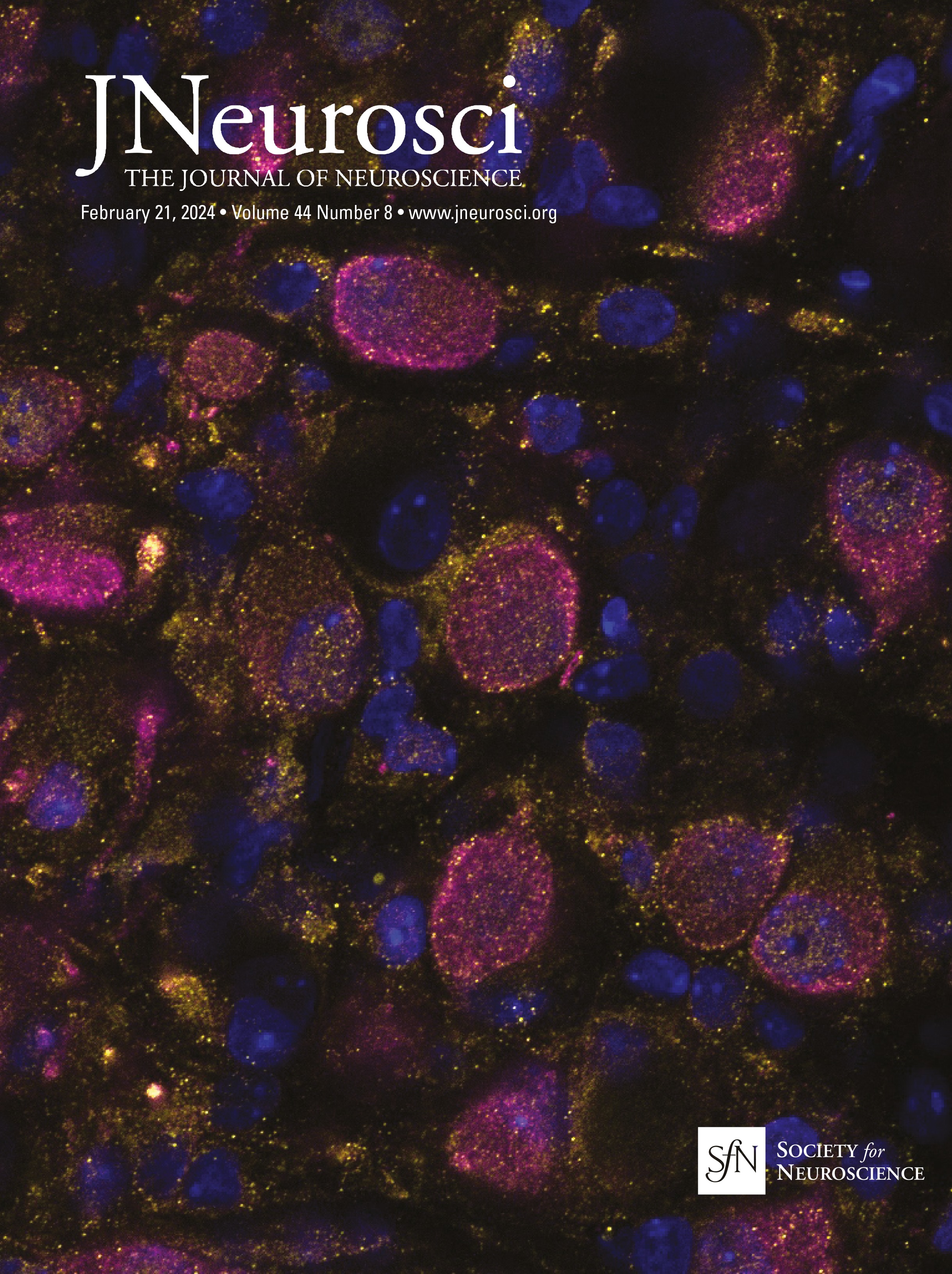
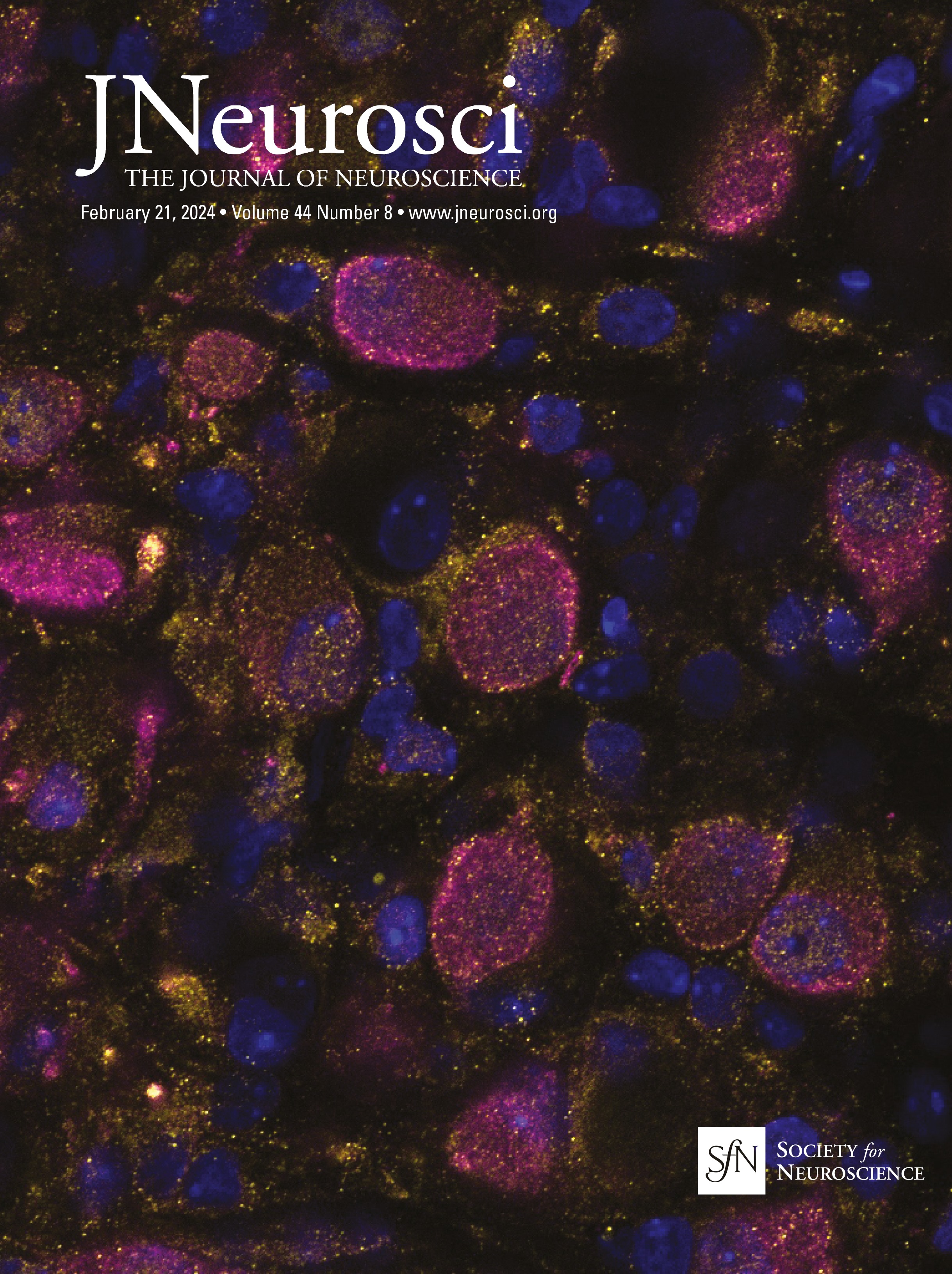
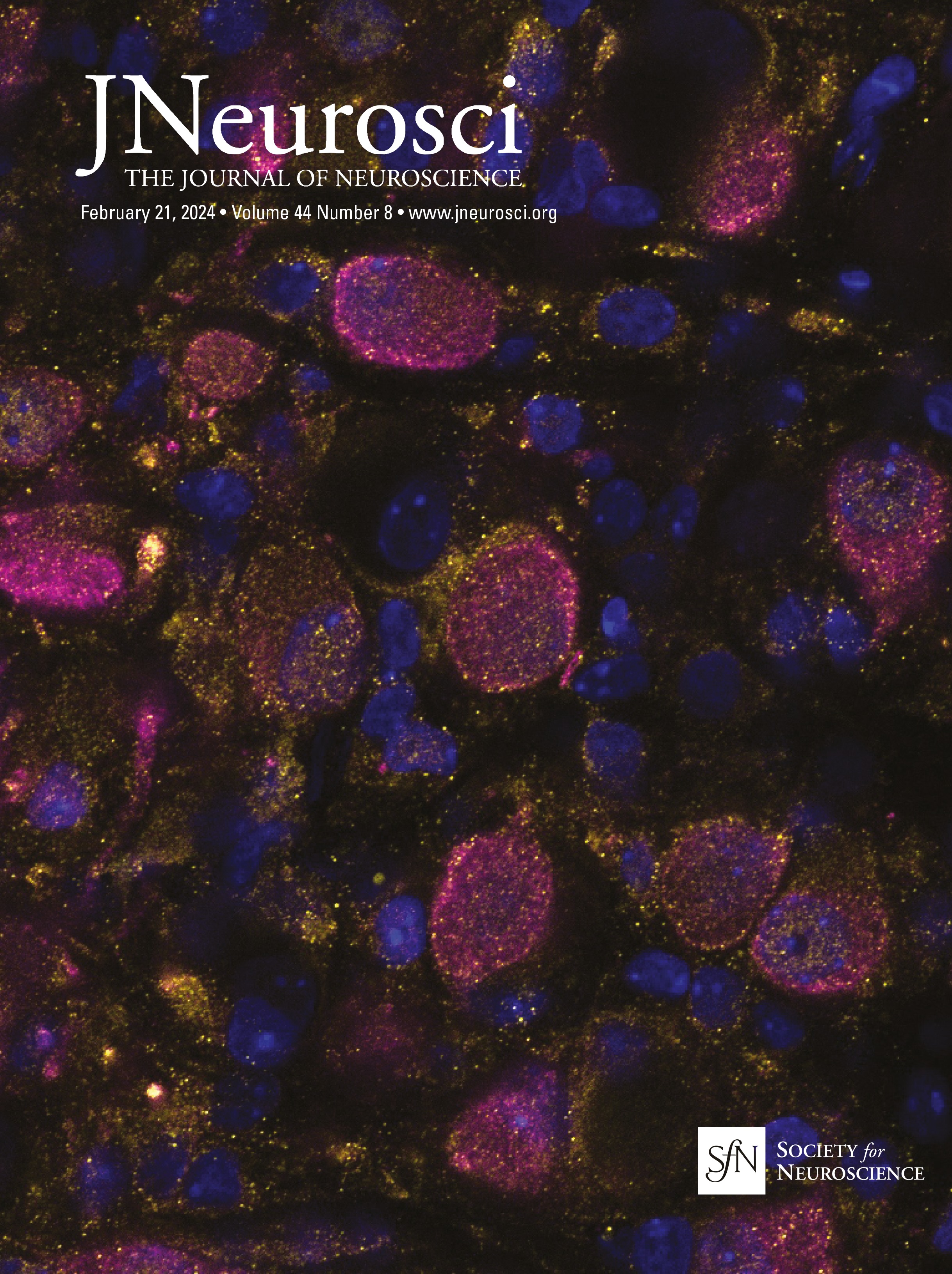
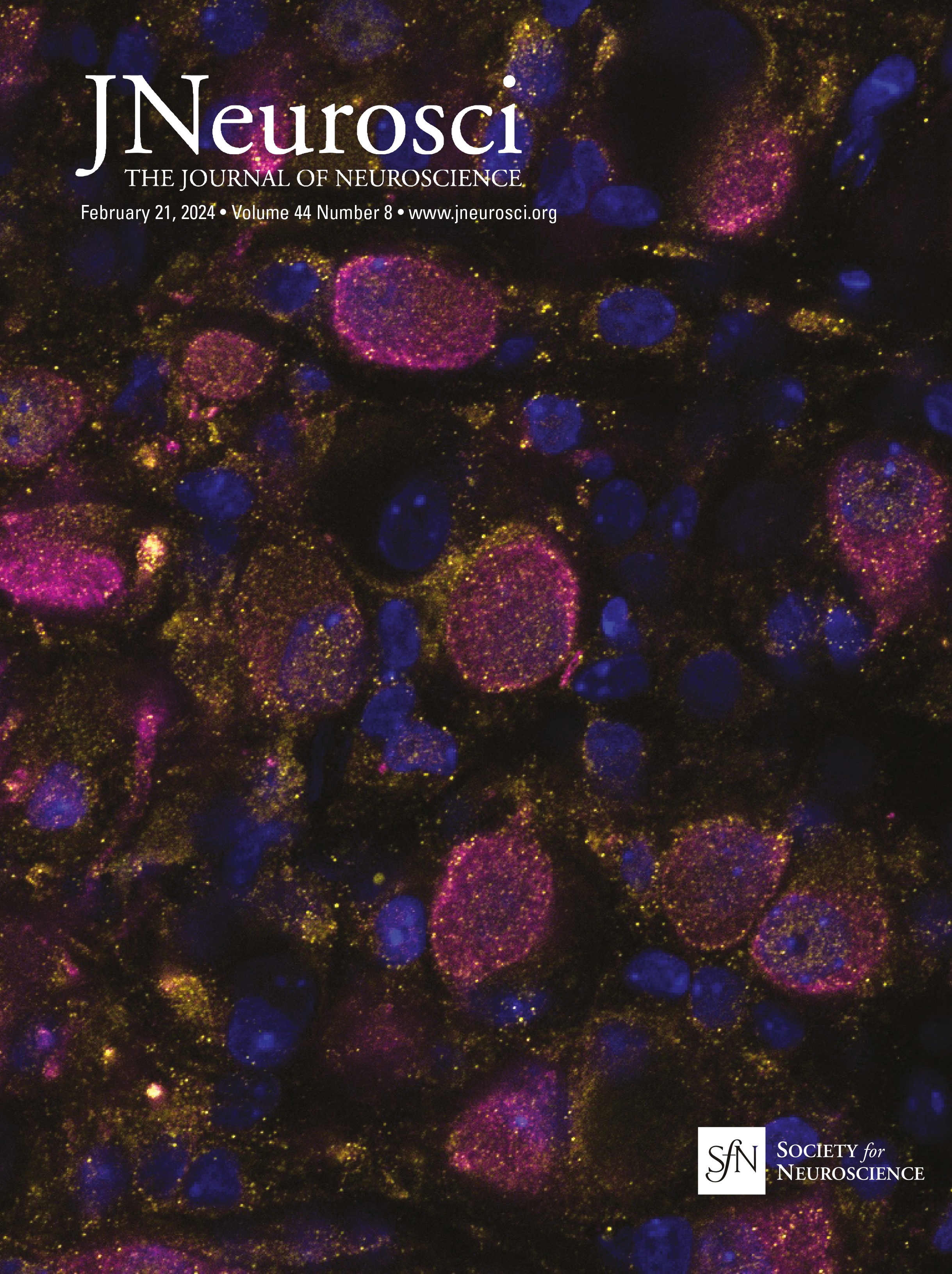
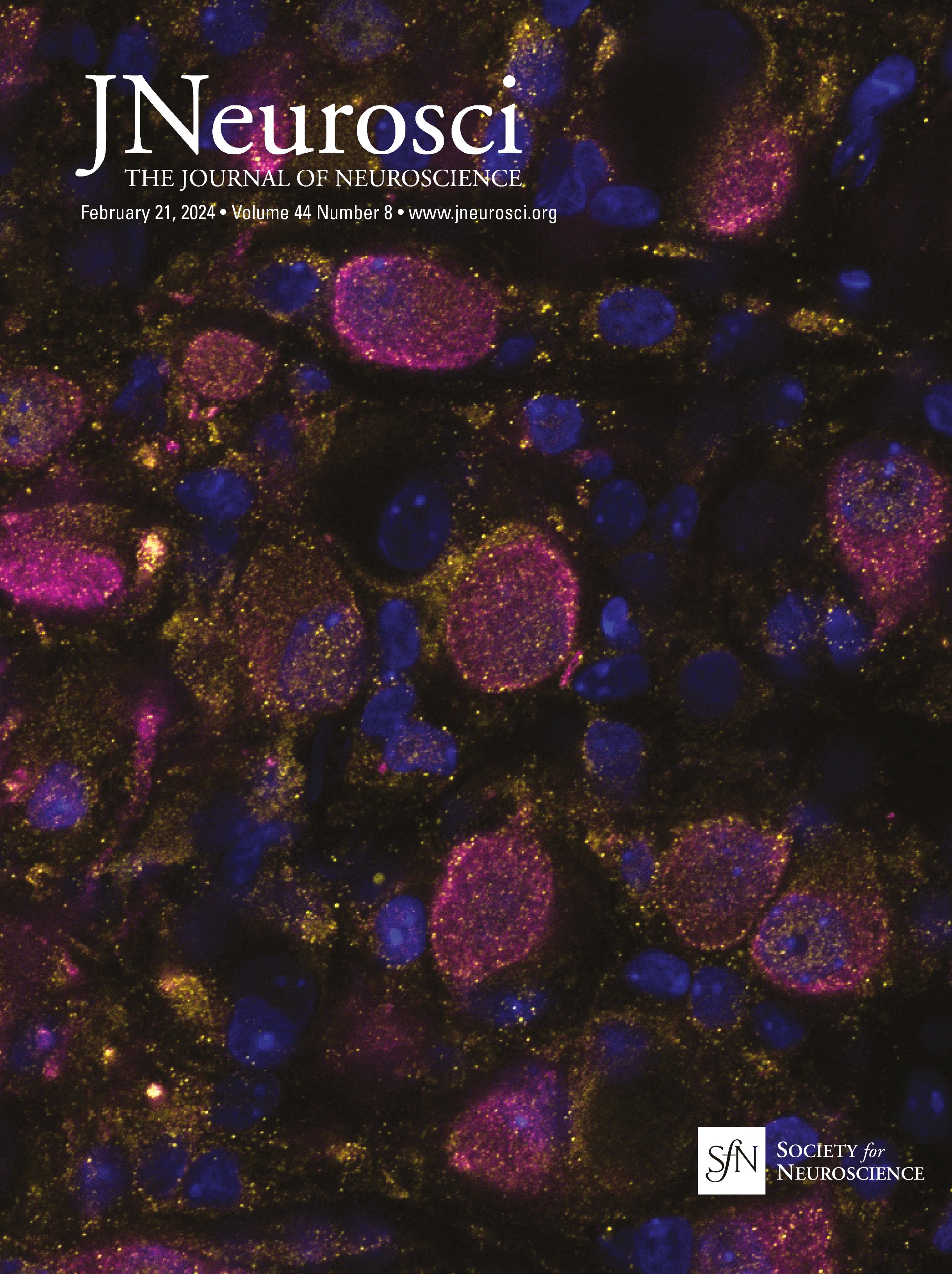
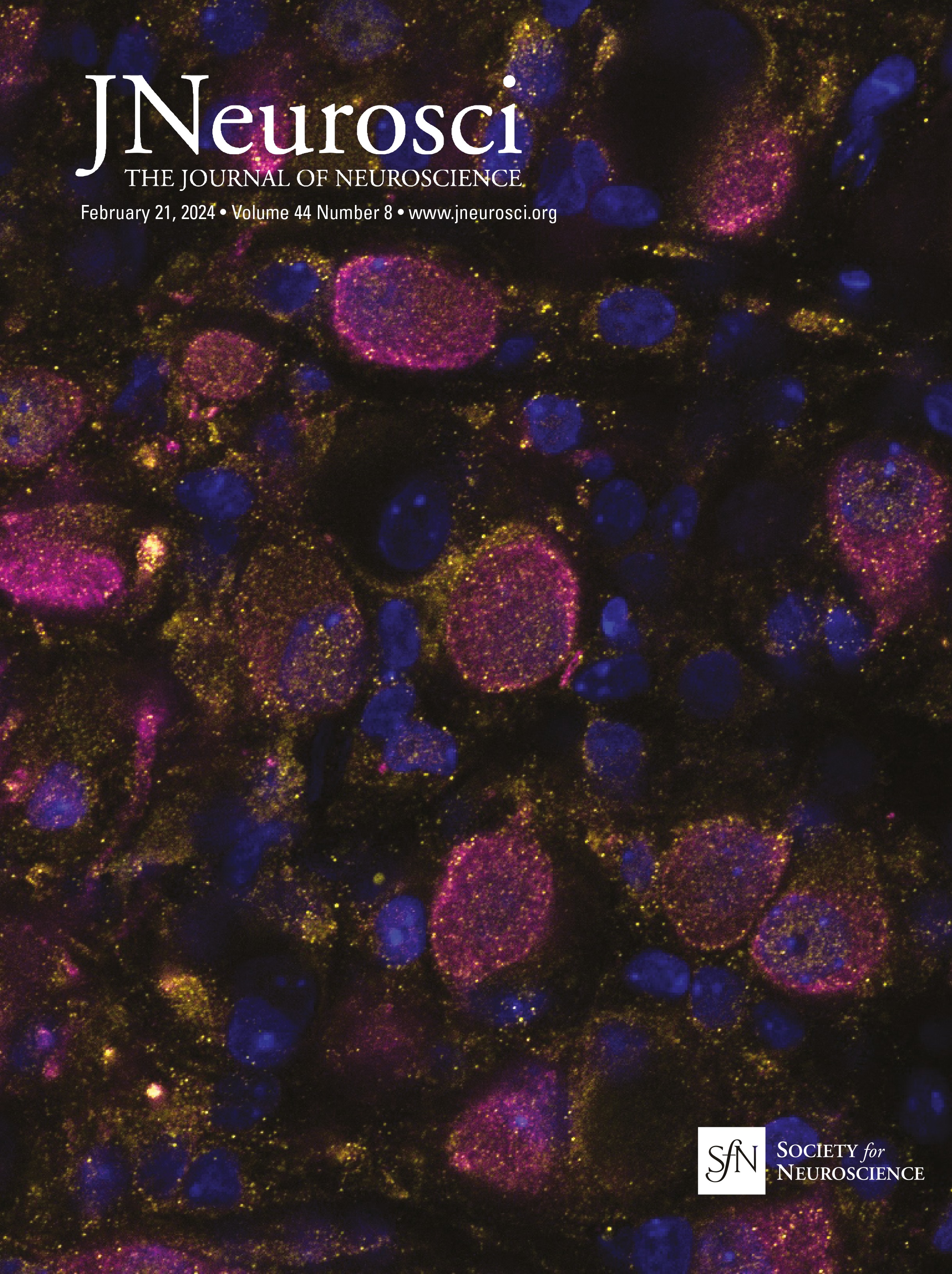
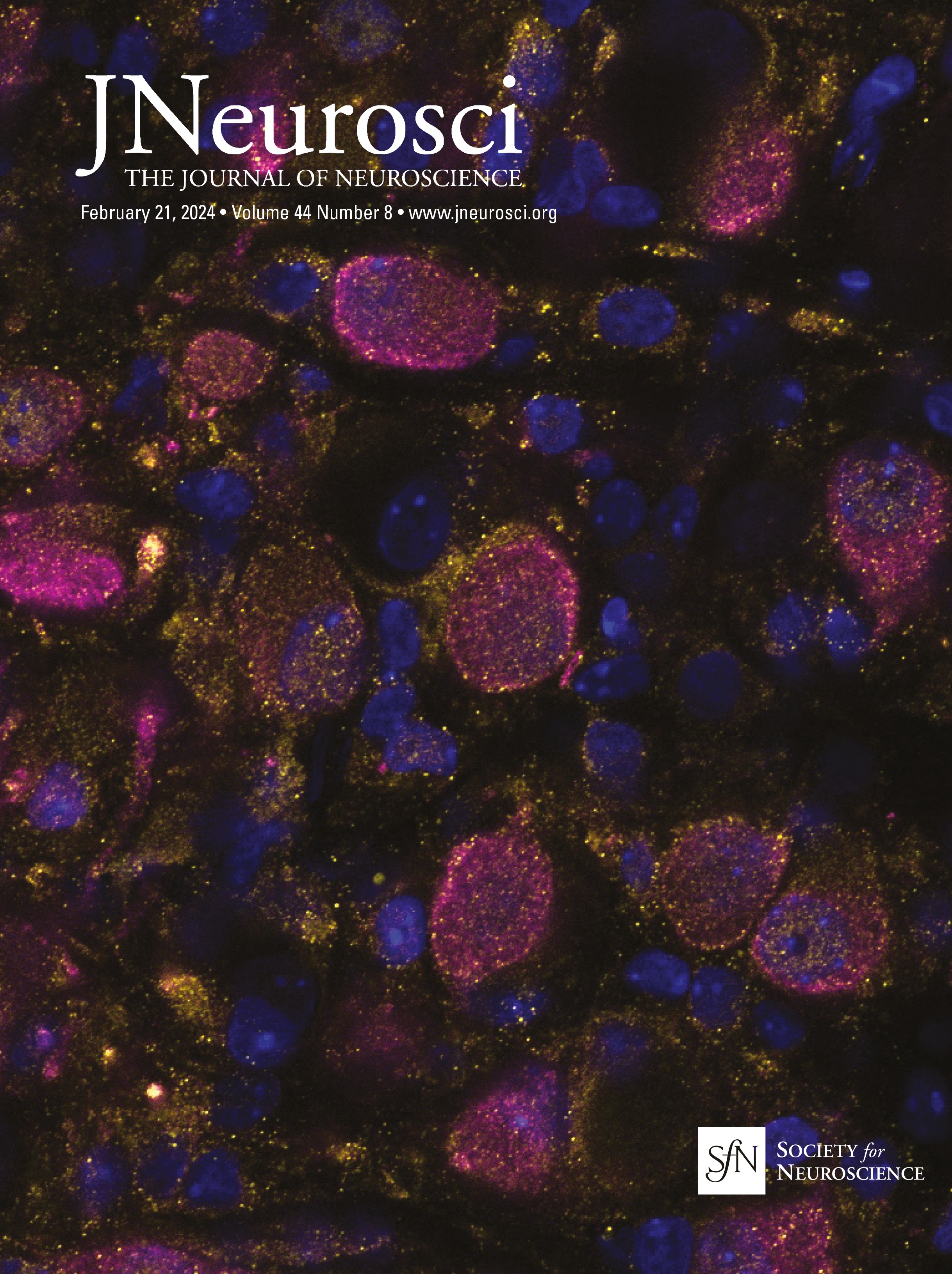
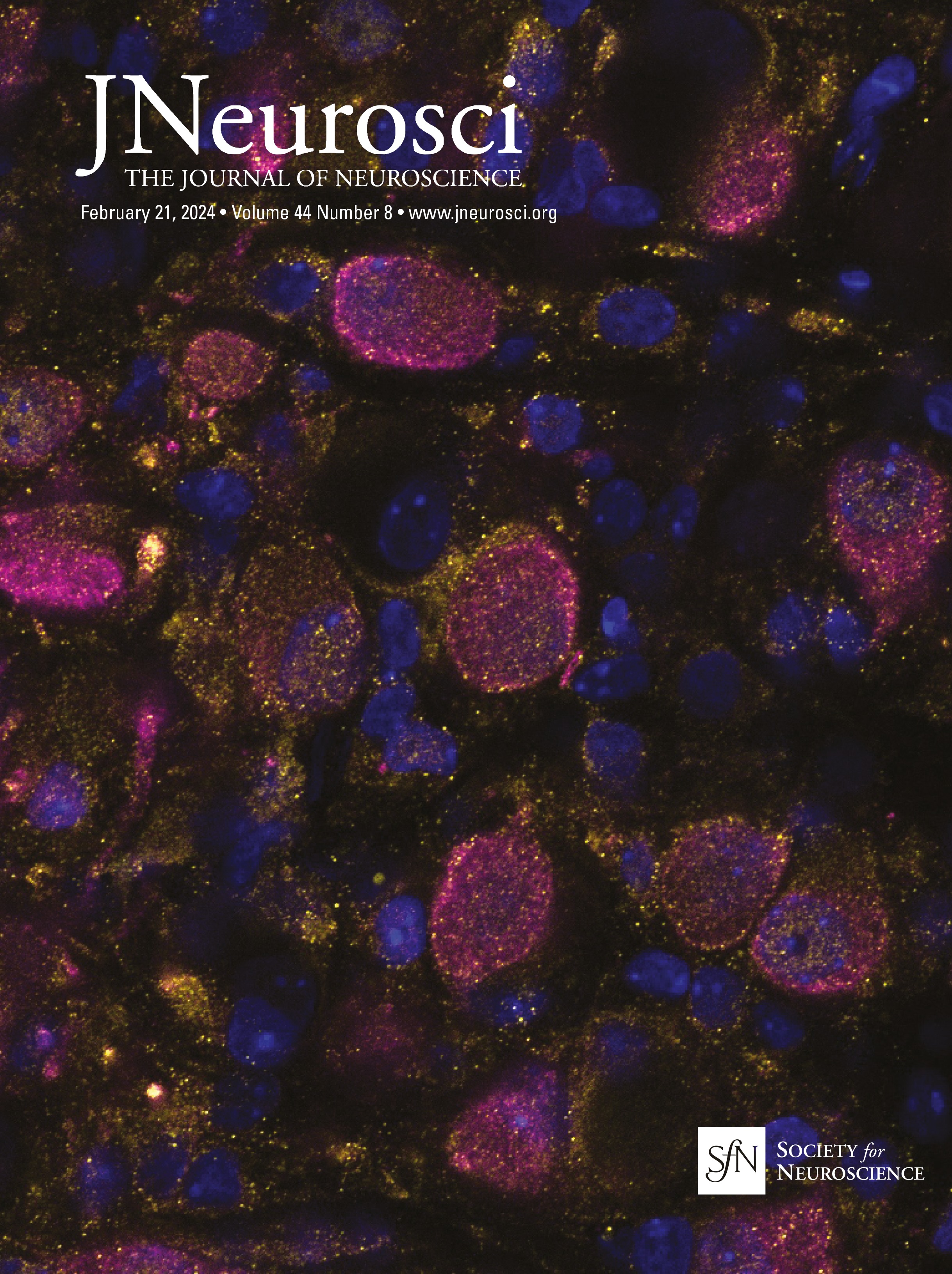
Figure 1. IO neurons can be divided into distinct morpho-anatomical types. A, Dorsal and lateral views of the average expression of the hspGFFDMC28C line used in this study (green, N = 39 fish; line from Takeuchi et al., 2015) registered to a common reference larval zebrafish brain (gray), showing strong signal in the IO and in the CFs in the cerebellum. In this and subsequent panels: ro, rostral direction; l, left; r, right; c, caudal; d, dorsal; v, ventral; scale bars, 100 µm; vertical dashed lines indicate the midline of the brain. Teal rectangle outlines the area shown in B, C, D, and F. B, Example of a single labeled IO neuron (magenta). C, Axon reconstruction of that neuron. The inset shows its dendritic morphology, and the asterisk indicates its axon. D, Axon reconstruction of all labeled IO neurons (N = 53 neurons from 39 larvae) color coded by soma location (red, left IO, teal, right IO), showing that IO neurons project contralaterally. E, Examples of IO neurons that were divided into two morphological classes: unipolar neurons (green) that have a single dendritic tree arborized along the midline, and multipolar neurons (magenta) that have bi- or tri-polar dendritic trees. Asterisks indicate axons. For the complete dataset (N = 16 unipolar, 19 multipolar and 18 ambiguous neurons) see Figure 1-1. F, Axon reconstruction of all unipolar (green) and multipolar neurons (magenta), showing that the morphological type of a neuron is predictive of its projection pattern and its location within the IO.
Figure 1-1The complete dataset of single-labeled IO neurons A. Dendritic morphology of all labeled IO neurons divided by their morphological class (green, 16 unipolar neurons, magenta, 19 multipolar neurons, black, 18 ambiguous neurons). Asterisks indicate axons. B. Axon reconstruction of all labeled neurons; ro, rostral direction, l, left, r, right, c, caudal, d, dorsal, v, ventral; scale bars, 100 µm. Download Figure 1-1, TIF file.
 Figure 2.
Figure 2. The majority of IO neurons are sensitive to translational and rotational motion, direction-selective and spatially organized. A, Left, two-photon laser-scanning microscope (2P LSM) used for calcium imaging. Middle, same as Figure 1A; ro, rostral direction; l, left; r, right; c, caudal; scale bars, 100 µm. Teal rectangle outlines the imaged area. Right, maximum intensity z-projection of the anatomy stack from an example fish, showing a typical imaging field of view with individual IO neurons. B, Stimulation protocol. Fish were presented with translational and rotational motion, with each stimulus lasting 21.4 s (6 s stationary, 10 s moving, 5.4 s stationary). Translational gratings moved at 10 mm/s and rotational windmill at 22.5 °/s. For each imaging plane, we presented translational gratings in 8 different directions in a randomized order, followed by clockwise (CW) and counter-clockwise (CCW) rotational motion. C, Average responses of four forward-selective example neurons to translational and rotational motion. DS, direction-selective neurons responding exclusively to translation motion, DS + CW or DS + CCW, direction-selective neurons that also responded to CW or CCW rotation, DS + CW + CCW, direction-selective neurons that also responded to rotation in both directions. Shadows represent SEM across repetitions. Horizontal dotted lines represent each neuron baseline. In C and D, vertical dotted lines separate stationary and moving periods of the stimulus, vertical solid lines separate visual stimuli. D, Left, raster plot for all direction-selective neurons (N = 608 neurons from 12 fish), grouped by response types and sorted by PD (coded by the circular color wheel). Each row represents a neuron’s average response to the ten stimuli shown on top. Right, average response of all neurons in each group to CW and CCW stimuli. Shadows represent SEM across neurons. E, Left, distribution of PDs of all direction-selective neurons and of the 4 different groups of neurons independently. 0° represents forward, 90° rightward, 180° backward and −90° leftward directions. Right, probability distribution of the cosine of PD for each of the four groups. F, Spatial distribution of PDs within the IO (N = 967 active neurons, including 608 direction-selective ones, from 12 fish). Each dot represents a neuron color-coded for PD as represented in the color wheel, or gray for not direction-selective neurons. Small italic roman numbers indicate the location of neurons shown in C. In F and G: ro, rostral direction; l, left; r, right; c, caudal; scale bars, 25 µm. Green and magenta curves show the rostral–caudal distribution of forward- and backward-preferring neurons respectively, separated according to the sign of the PD cosine. Blue and orange curves show the left-right distribution of left- and right-preferring neurons respectively, separated into forward-preferring (top) and backward-preferring (bottom) groups. G, Spatial distribution of rotation sensitivity within the IO. Each dot represents a neuron color-coded for its response to CW (red) and CCW (blue) stimuli. White lines on the color bars denote 99th and 95th percentiles, used for presenting the spatial distributions of top 1%, top 5%, and remaining 95% active neurons.
 Figure 3.
Figure 3. The majority of IO neurons receive input from both eyes with a contralateral bias. A, Monocular visual stimuli. Fish were presented with translational and rotational motion, with each stimulus lasting 21.4 s (6 s stationary, 10 s moving, 5.4 s stationary). Translational gratings moved at 10 mm/s and rotational windmill at 22.5°/s. Each stimulus was presented three times per imaging plane: to the left eye, to the right eye, and binocularly. To avoid contralateral contamination during monocular stimulation, the two visual fields were separated by a vertical 0.5 mm black patch that was positioned below the fish body. In addition, we had a 55° cut-off in front of the fish (27.5° in each eye) to prevent stimulation of the eyes’ binocular zone. B, The trial structure was the same as in the binocular stimulation experiment (translational gratings in 8 different directions in a randomized order, followed by clockwise (CW) and counter-clockwise (CCW) rotational motion). To minimize possible contribution of light onset/offset to the responses, monocular and binocular stimulation were performed in groups: left, followed by right and finally binocular stimulation. Binocular stimulation block also included converging (conv) and diverging (div) rotational motion. C, Examples of neurons’ responses to monocular stimulation of the left (cyan) and right (yellow) visual fields and to binocular stimulation (magenta). Vertical dotted lines separate stationary and moving periods of the stimulus. Horizontal dotted lines represent each neuron baseline. Vertical solid lines separate different visual stimuli. For comparison purposes, all example neurons monocular CW and CCW responses are repeated in binocular convergence and divergence stimuli (dashed lines). Shadows represent SEM across repetitions. D, Distribution of neurons’ monocular index. Monocular bias is color coded with a red (contra) through gray (binocular) to blue (ipsi) gradient. Dashed line indicates unbiased binocular neurons. Note that the distribution is shifted to the right, indicating that IO neurons are in general more sensitive to contralateral stimulation. E, Spatial distribution of monocular bias within the IO, color coded as histogram in D; ro, rostral direction; l, left; r, right; c, caudal; scale bar, 25 µm. Small italic roman numbers indicate the location of neurons shown in C. N = 518 neurons (of which 511 were active) from six fish.
Figure 3-1Rotation-sensitive IO neurons often receive inputs with opposing PDs and occupy distinct regions within the IO A. Left, maximal response to rotational stimuli as a function of the absolute monocular index of each neuron (left) and of absolute difference in the PDs of the left and right visual fields (right). B. Binocular PD as a function of left and right eye PDs, color-coded as in color wheel on top. Gray dots represent non direction-selective cells. In B, C, and D: small italic roman numbers indicate example neurons shown in Figure 23. C. Responses to binocular CW rotation (left) and CCW rotation (right) as a function of left and right PDs. D. Spatial organization of the IO neurons, color-coded based on their monocular PDs. Shown in red and blue are neurons with opposing PDs (upper left and bottom right quadrants in B and C). Green and magenta indicate neurons with similar PDs (green: forward monocular PDs, bottom left quadrant; magenta: backward PDs, upper right quadrants). Ro, rostral direction, l, left, r, right, c, caudal; scale bar, 25 µm. Small italic roman numbers indicate the location of neurons shown in C. N = 518 neurons (of which 511 were active) from 6 fish. Download Figure 3-1, TIF file.
 Figure 4.
Figure 4. Functional organization of the IO maps onto its morpho-anatomical organization. A, Left, light-sheet microscope used for fast volumetric calcium imaging. Right, same as Figure 1A. Teal rectangle outlines the imaged area. B, Stimulation protocol. The trial structure was the same as in the binocular stimulation experiment [translational gratings in 8 different directions in a randomized order, followed by clockwise (CW) and counter-clockwise (CCW) rotational motion]. Additionally, we included converging (conv) and diverging (div) rotational motion. Each stimulus lasted 21 s (6 s stationary, 10 s moving, 5 s stationary). Translational gratings moved at 10 mm/s and rotational windmill at 22.5 °/s. For each fish, we presented this stimulus set five times. C, Max z-projection of the distribution of active voxels categorized as forward selective (green), backward selective (blue) or rotation selective (red), averaged across fish. D i, Max z-projections of the distribution of active voxels categorized as left selective (green) or right selective (magenta) in the entire imaging field of view. D ii, Max lateral projection of the distribution of active voxels selective for ipsiversive motion (green) and for contraversive motion (magenta) within the IO. E i, Max z-projection patterns of unipolar and multipolar neurons from Figure 1F, color-coded depending on their morphological type and left-right location within the IO. E ii, Lateral projection of the location of unipolar (green) and multipolar neurons (magenta) within the IO. F, Overlay of D and E. Note that the distribution of active voxels in the IO in the light-sheet imaging data includes signals not only from cell somata but also from surrounding neuropil, which accounts for the more lateral spread compared to the soma distribution. See Table 1 for quantification of the overlap. N = 28 fish; ro, rostral direction; l, left; r, right; c, caudal; v, ventral; d, dorsal; scale bars, 100 µm.
Figure 4-1Functional mapping based on responses to rotational, forward and backward motion does not correspond to the morpho-anatomical organization of the IO A. Max projections of the distribution of active voxels color-coded using different functional axes: A i, rotational-selective (green) VS backward-selective voxels (magenta); A ii, forward-selective (green) VS backward-selective voxels (magenta). A iii is reproduced from Figure 4D for comparison to other functional mappings. Images on top show max z-projections of the entire imaging field of view that included IO and CFs, bottom images show zoomed-in lateral projections zoomed into the IO. B. Overlap of respective functional maps with morpho-anatomical mapping within the IO. Each dot represents one unipolar or multipolar neuron from Figure 1. See Table 1 for quantification of the overlap. N = 28 fish; ro, rostral direction, l, left, r, right, c, caudal, v, ventral, d, dorsal; scale bars, 100 µm. Download Figure 4-1, TIF file.
Experiments 1–3 allowed us to formulate three specific hypotheses for how functional organization might be related to morpho-anatomical organization. In Experiment 4, we specifically test these hypotheses using a bootstrapping procedure described in the Materials and Methods section, Quantification of overlap (see below). The significance level was set to 5% and p-value thresholds were Bonferroni-corrected for six comparisons (3 hypotheses times two brain regions: IO and cerebellum). We used one-tailed alternatives because results of Experiments 1–3 allowed us to generate specific expectations of the relationship between function and anatomy.
Experiments were performed in accordance with the European Directive 2010/63/EU and approved by the Champalimaud Ethics Committee and the Portuguese Direcção Geral Veterinária (Ref. No. 019774) and approved protocols set by the Max Planck Society and the Regierung von Oberbayern (TVA 55-2-1-54-2532-82-2016).
All data and code used in this study can be made available upon request.
Experimental animalsZebrafish husbandryAll experiments were performed on larval zebrafish (Danio rerio) at 6–7 d post-fertilization (dpf), with the exception of single-cell electroporation (see below) that was performed at 5–6 dpf. The sex of the animals could not be determined at this early developmental stage.
Zebrafish breeding and maintenance were performed under standard conditions (Westerfield, 2007; Martins et al., 2016). Both adult fish and larvae were maintained at 28°C on a 14/10 h light/dark cycle. Adult zebrafish were housed in a zebrafish facility system with constantly recirculating water with about 10% daily water exchange. Fertilized embryos were collected in the morning and kept in 94 mm Petri dishes at a density of 20 animals per dish in E3 medium with daily water exchange, unless otherwise specified.
Transgenic linesAll experiments were performed using a transgenic hspGFFDMC28C driver line (rk8Tg; Takeuchi et al., 2015), expressing a modified version of Gal4 mainly in the IO neurons (Fig. 1A). Animals were also homozygous for the nacre mutation, which introduces a deficiency in the mitfa gene involved in development of skin melanophores (Lister et al., 1999), thereby allowing for noninvasive brain imaging. The UAS reporter gene differed depending on the experiment.
For single-cell electroporation, we used the incross of hspGFFDMC28C; UAS:mCherry to inject plasmid DNA pCS2-GAP43-GFP construct (kindly provided by Isaac Bianco).
For sparse single-cell genetic labeling, we outcrossed hspGFFDMC28C; UAS:GFP to UAS:epNtr-tagRFP reporter line (line mpn123 generated by Miguel Fernandes and Herwig Baier at MPI Neurobiology).
For functional imaging experiments, we used the offspring of an incross of hspGFFDMC28C; UAS:GCaMP6fEF05 (ccu2Tg) for IO-specific expression of the calcium indicator GCaMP6fEF05. This modified version of GCaMP6f (Chen et al., 2013) was made by making the mutations D397N/G398A/N399D (Sun et al., 2013) in the CaM domain of GCaMP6f. This version reports activity in zebrafish neurons with better signal to noise ratio than GCaMP6f, while maintaining its fast dynamics in comparison to GCaMP6 s (Ostrovsky, Renninger et al., in prep.).
Experiment 1: morpho-anatomical characterization of the IO neuronsThe first aim of this study was to characterize the morpho-anatomical properties of the IO neurons in larval zebrafish by the means of single-cell labeling (Fig. 1). Labeling of individual IO cells was achieved by either single-cell electroporation or by sparse genetic labeling.
Single-cell electroporation5–6 dpf hspGFFDMC28C; UAS:mCherry larvae were embedded in 1.5% low melting point agarose, anesthetized in bath-applied solution of MS-222 (tricaine) at a concentration of 0.16 g/L in Danieau’s solution (58 mM NaCl, 0.7 mM KCl, 0.4 mM MgSO4, 0.6 mM Ca(NO3)2, 5 mM HEPES buffer), and IO neurons were electroporated under a confocal microscope (LSM 780, Carl Zeiss) as described previously (Tawk et al., 2009). Briefly, a fine borosilicate glass electrode with filament (final tip diameter ∼1 µm) was filled with plasmid DNA pCS2-GAP43-GFP construct (kindly provided by Isaac Bianco) at a concentration of ∼1 µg/µl in distilled water and manipulated through the tissue to a target mCherry-positive IO neuron using a micromanipulator (Sutter Instruments). 1–3 square trains of electric pulses with a frequency of 200 Hz, duration of 1 s, and magnitude of 20–30 V were applied to inject DNA constructs into a single neuron using an Axoporator 800A (Molecular Devices). After electroporation, dishes with embedded larvae were gently washed with Danieau’s solution three times to wash out the tricaine. After washing, larvae were released from the agarose and allowed to recover in Danieau’s solution overnight.
Sparse genetic labelingSparse genetic labeling of individual IO neurons was achieved by outcrossing the hspGFFDMC28C; UAS:GFP to a UAS:epNtr-tagRFP reporter line. The offspring of such an outcross typically had very sparse expression of RFP, often in only one or two IO neurons, which was ideal for neuronal tracing.
Confocal imagingTo image the labeling results, 6–7 dpf larvae were anesthetized using tricaine and embedded in 1.5% low melting point agarose. Labeled neurons were imaged using a confocal microscope (LSM 780, Carl Zeiss). For each successfully labeled larva, two z-stacks were acquired: one for visualizing the cell body and dendritic arbors at higher magnification (Fig. 1E and Fig. 1-1A), and another to capture the whole span of axonal projections at lower magnification. The second stack was acquired in two channels: a single-cell channel (GFP for the electroporated larvae and RFP for larvae with sparse genetic labeling) and an anatomical reference channel containing dense signal from the majority of the IO neurons and their projections (mCherry and GFP, respectively) (Fig. 1B). The latter channel was used for anatomical registration of the data to a standard reference brain (see Anatomical registration).
TracingAfter anatomical registration, axonal projections of labeled IO neurons were traced and reconstructed using the “Simple Neurite Tracer” FIJI plugin (Longair et al., 2011; Schindelin et al., 2012) (Fig. 1C). All animals, where IO morphology and projection pattern could not be clearly and unambiguously traced (due to too dense labeling or low signal), were not analyzed. After exclusion, 39 animals were used to label 53 individual IO neurons.
Experiment 2: responses to whole-field visual stimulationThe next aim was to characterize the activity of the IO neurons in response to whole-field translational and rotational visual motion using two-photon calcium imaging in awake animals (Fig. 2). In total, we recorded activity of 1,106 IO neurons from 12 larvae expressing an enhanced version of GCaMP6f specifically in the IO neurons (see Transgenic lines for more detail).
Two-photon functional imagingFunctional imaging experiments were conducted on head-restrained preparations of 6–7 dpf zebrafish larvae (Portugues and Engert, 2011). Each larva was embedded in 2% low melting point agarose (Invitrogen, Thermo Fisher Scientific) in a 35 mm Petri dish with a Sylgard 184 base (Dow Corning). After allowing the agarose to set, the dish was filled with E3 medium, and the agarose around the tail and eyes was removed to allow for tail and eye movements that were used as a readout of behavior. Note that, in this study, the recorded behavioral traces were not analyzed.
The dish with the embedded larva was then placed onto a light-diffusing screen, located 5 mm below the larva, and imaged on a custom-built two-photon microscope (Fig. 2A, left).
A Ti-Sapphire laser (Coherent Chameleon) tuned to 950 nm wavelength was used for excitation and custom-written Labview software was used to control the microscope and to capture image data. Larval brains were systematically imaged from dorsal to ventral, in 2 µm z-steps, at approximately 3 Hz (345.6 ms/frame). In each fish, we imaged 30–40 planes which corresponded to 60–80 µm that covered the entire IO volume (Fig. 2A, right).
In addition, two infrared LEDs (850 nm wavelength) were angled between the imaging objective and the petri dish to illuminate the fish and allow the tracking of the tail. Behavior was recorded using a Mikrotron EoSens (MC1362) high-speed camera and a National Instruments frame grabber (PCIe-1433) (not shown in Fig. 2A). Tail tracking was performed at 700 Hz and eye-tracking at 100 Hz using a custom-written software in C#.
Binocular visual stimuliFor visual stimulation, we used a custom-written rendering engine that uses fragment shaders in OpenGL to draw visual stimuli in synchronization with two-photon imaging software. Visual stimuli were projected from below onto a flat screen at 60 Hz using a Optoma ML750e LED projector and a red colored glass long-pass filter (Thorlabs FGL590) and Texas Red bandpass emission filter (Thorlabs MF630-69) to allow for simultaneous imaging and visual stimulation.
Visual stimuli were projected from below onto a flat diffusing screen, located 5 mm below the fish and centered under its head. The stimulus set consisted of 10 stimuli per imaging plane: whole-field sine black-and-red gratings with a 10 mm spatial period moving in eight directions at 10 mm/s in a randomized order, followed by whole-field square black-and-red windmill stimulus rotating at 22.5°/s clockwise (CW) and then counterclockwise (CCW) (Fig. 2B). Each stimulus lasted 21.4 s (6 s stationary, 10 s moving, 5.4 s stationary), which corresponded to 62 imaging frames. After presenting the complete stimulus set, the imaging plane was moved 2 µm ventrally and the set of stimuli was repeated.
In our study, we show visual stimuli on a screen below the fish, as is the common practice in larval zebrafish. These stimuli have been well characterized in the context of optomotor and optokinetic behavior but do not allow mapping of visual responses originating from the upper visual field, in contrast to studies in other species which have used planetarium style surround stimulation. In a recent study in zebrafish, motion noise stimuli were presented on a vertical cylindrical screen to map the spatial receptive fields of optic flow responses in the pretectum of the zebrafish larva (Zhang et al., 2022). It was found that, of the neurons with bimodal receptive fields that could distinguish translational and rotational motion, a large majority was sensitive to translation in the horizontal plane, and horizontal rotations, consistent with the stimuli used in our study.
Image registrationImage processing and analysis was performed automatically using custom-written MATLAB code (MathWorks). To correct for motion artifacts and possible drifting of the animals inside the agarose, acquired frames were first aligned within a plane then across planes similarly to (Portugues et al., 2014). Any experiments during which the fish drifted significantly in z were stopped and the data discarded. If a frame could not be aligned to adjacent ones due to strong movement artifacts, this frame was excluded from the analysis. To generate anatomy stacks for each larva, all aligned frames within each plane were averaged. We then registered the anatomy stacks of individual fish to
留言 (0)