
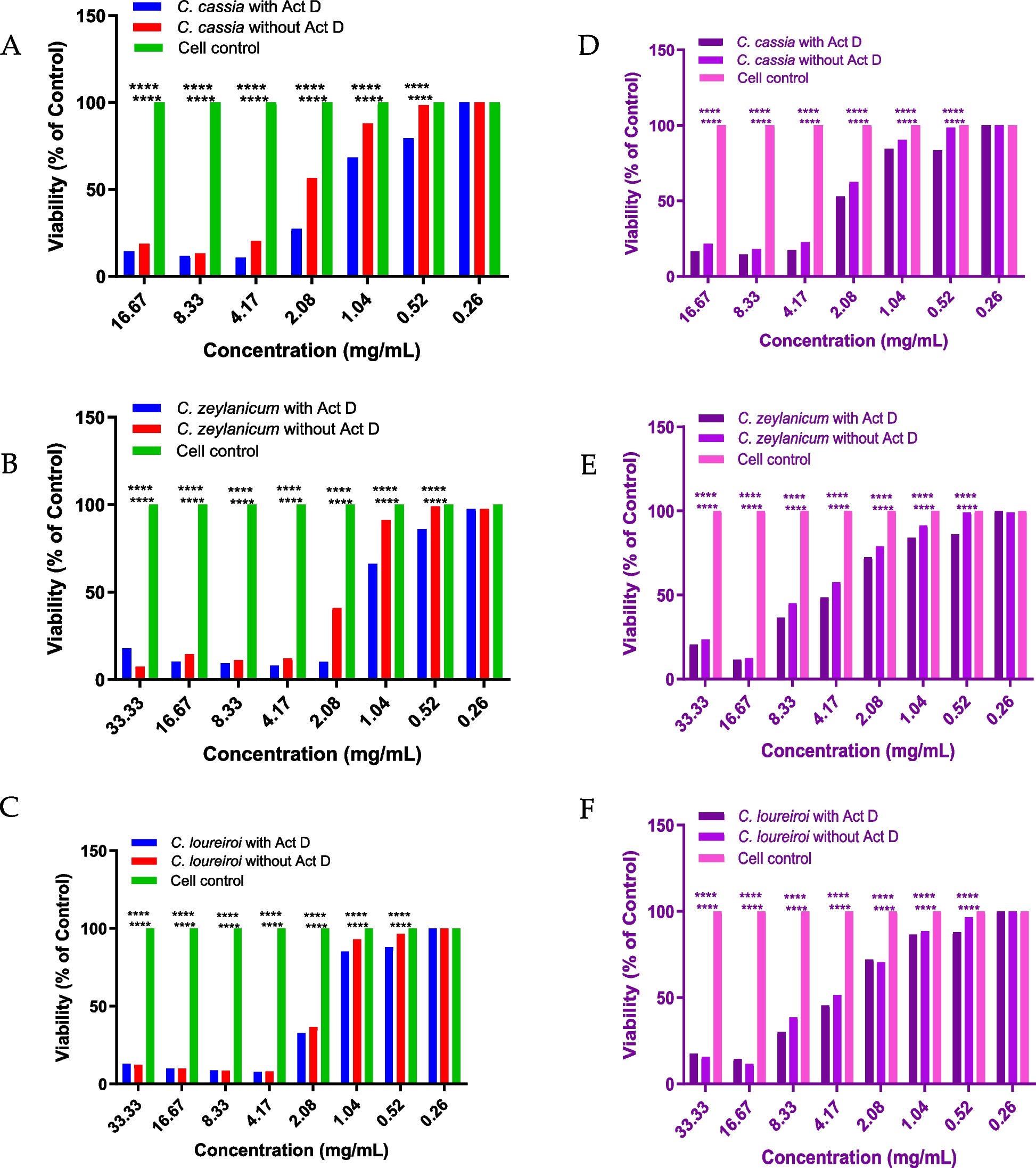
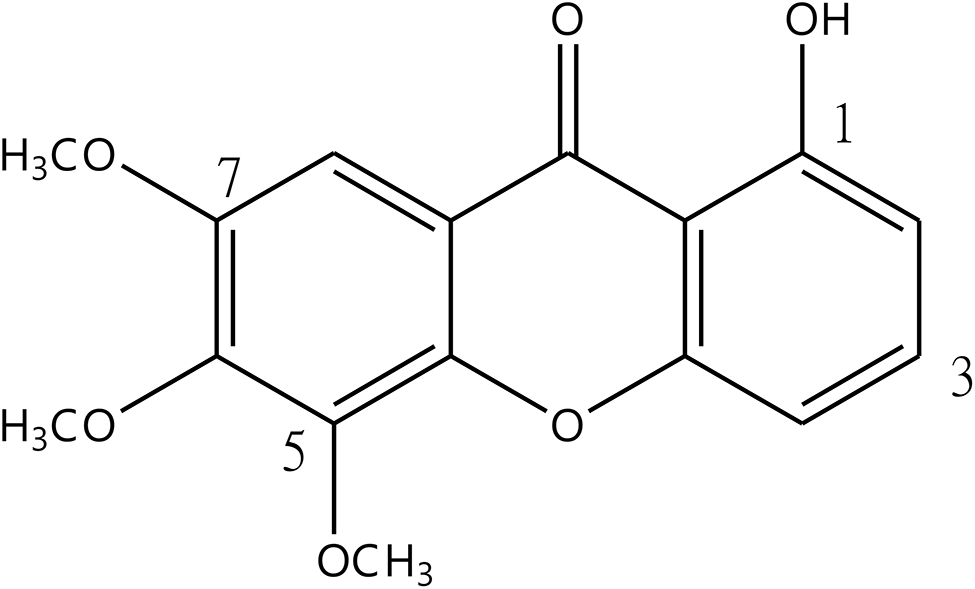
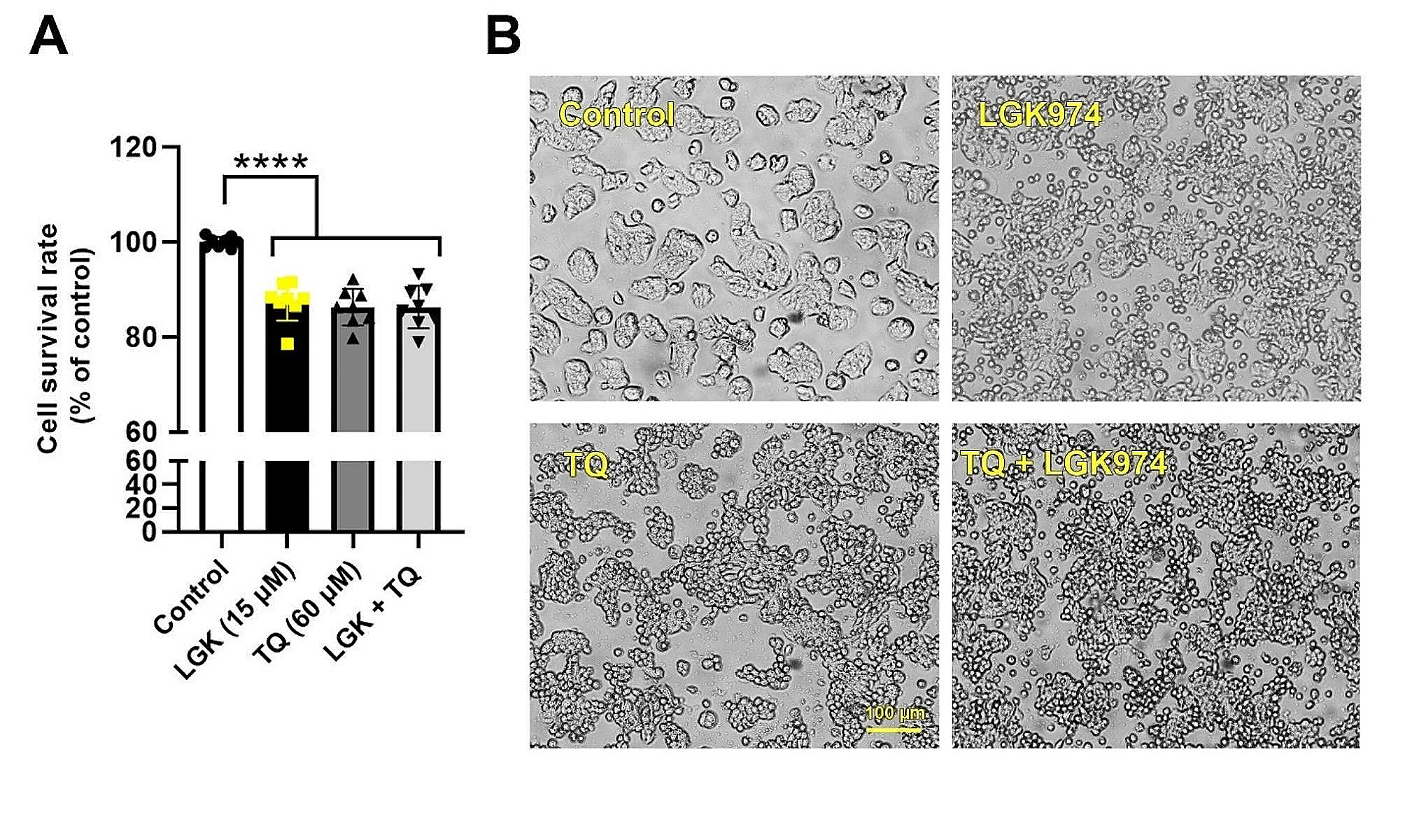
記住我






To explore the anticancer activity of diosbulbin C in NSCLC, a series of cytological experiments was conducted. Firstly, the impact of diosbulbin C on cell viability was investigated using the CCK-8 kit. After exposure to different concentrations of diosbulbin C for 48 h, dose-dependent suppression of diosbulbin C on cell viability was observed in NSCLC cells (Fig. 2A and Supplementary Figure S1). The IC50 (half-maximal inhibitory concentration) values of 100.2 μM, 141.9 μM, and 228.6 μM in A549, H1299, and normal lung HELF cells, respectively, were obtained, suggesting the relatively low cytotoxicity of diosbulbin C to normal cells. Besides, diosbulbin C treatment resulted in significant suppression of NSCLC cell proliferation as observed under the microscope (Fig. 2B). The inhibitory effect of diosbulbin C on the proliferation of NSCLC cells was further evaluated by colony formation and EdU assays. The colony formation results showed that diosbulbin C significantly inhibited the colony formation of NSCLC cells (Fig. 2C). And compared to the control group, the percentage of EdU-positive cells in diosbulbin C treatment groups was also significantly decreased (Fig. 2D). These findings demonstrate the anti-proliferative effect of diosbulbin C on NSCLC cells.
Fig. 2
Diosbulbin C inhibits cell viability and proliferation in NSCLC cells. A Cell viability was detected using CCK-8 assay after treatment with different concentrations of diosbulbin C in A549, H1299, and HELF cells for 48 h. B The cell morphology change was detected under a microscope after the treatment with diosbulbin C for 48 h. C Results of the colony formation of A549 and H1299 cells after the treatment with different concentrations of diosbulbin C. D Representative results of EdU assay of A549 and H1299 cells with/without diosbulbin C treatment
To determine the effect of diosbulbin C on cell apoptosis, we carried out an Annexin V-FITC dual staining assay with flow cytometry. As a result, we found that diosbulbin C can induce NSCLC cell apoptosis, however, only under relatively high concentrations (Fig. 3A). This result suggests that apoptosis induction may not be the predominant effect of diosbulbin C in NSCLC cells. Given the tight connection of cell cycle progression in cancer cells with cell proliferation, we then explored the impact of diosbulbin C on cell cycle in NSCLC cells. After exposing A549 and H1299 cells to 100 μM, 200 μM, and 300 μM of diosbulbin C for 48 h, the cell cycle was subsequently analyzed by flow cytometry. As a result, a significantly increased proportion of G0/G1 phase cells was observed in cells treated with diosbulbin C compared to the control group (Figs. 3B). The results demonstrated that diosbulbin C treatment potently induces G0/G1 phase cell cycle arrest and suppresses cell proliferation in NSCLC cells. Besides, we observed a relatively low proportion of G0/G1 phase cells in 300 μM of diosbulbin C treated group compared to 200 μM treatment, possibly due to the influence of significant cell death induced by high concentration of diosbulbin C (300 μM) in H1299 cells.
Fig. 3
A Representative results of annexin V-FITC/PI staining after the treatment with diosbulbin C for 48 h. B Representative results of cell cycle after the treatment with diosbulbin C for 48 h
AKT1, DHFR, and TYMS were identified as potential targets of diosbulbin C in NSCLCBased on the molecular formula of the diosbulbin C (Figs. 4A), its 100 potential targets were obtained from the Swiss Target Prediction (Supplementary Table S1). DisGeNET database was then used to identify the potential therapeutic targets of diseases, which resulted in 2438 target genes in NSCLC (Supplementary Table S2). The collective targets of diosbulbin C and NSCLC were obtained by merging the target data using the jvenn online tool, generating 43 overlapping target genes (Fig. 4B). The obtained 43 collective targets were considered as the potential effective targets of diosbulbin C in NSCLC. To better show the interactions of diosbulbin C with its potential targets in NSCLC, we subsequently constructed a “drug-target” network. The interactions between diosbulbin C and the targets are shown in Fig. 4C, which presents 44 nodes and 43 edges in this network, suggesting the multiple interactions of diosbulbin C with the potential targets.
Fig. 4
Potential target prediction of diosbulbin C in NSCLC. A The molecular structure of diosbulbin C. B The collective targets of diosbulbin C and lung cancer were obtained by Venn diagrams. C “Drug-target” network was constructed by cytoscape
To further explore the biological functions of the 43 target genes regulated by diosbulbin C in NSCLC and gain deep insights into the roles of diosbulbin C in biological processes against NSCLC, the Reactome enrichment and GO analysis of the obtained 43 target genes were performed using DAVID database. The results of Reactome enrichment analysis showed that the target genes were significantly enriched in 66 Reactome pathways. The top 25 remarkable enriched pathways are shown in Fig. 5A and Table 1. They were highly related to activation of matrix metalloproteinases, collagen degradation, extra-nuclear estrogen signaling, degradation of the extracellular matrix, immune system regulation, and PI3K/AKT signaling in cancer. In particular, AKT1, DHFR, and TYMS were mapped to the pathway of the mitotic G1 phase and G1/S transition and are closely related to cell cycle progression. To discover the core mechanism of the anti-NSCLC effect of diosbulbin C, all pathways and targets were subsequently subject to constructing a “target-pathway” network. As shown in Fig. 5B, the network comprises 108 nodes (42 marks and 66 signaling pathways) and 442 edges, and different node colors indicate the various targets and regulation pathways. Additionally, according to the GO analysis shown in Fig. 5C, the targets of diosbulbin C were mainly enriched in negative regulation of apoptotic process (GO:0043066), signal transduction (GO:0007165), and protein autophosphorylation (GO:0046777), associated with enzyme binding (GO:0019899), and protein tyrosine kinase activity (GO:0004713), etc. The above results suggest that diosbulbin C may exert its anticancer effect by directly targeting multiple targets associated with NSCLC and regulating the pathways involved in the biological processes.
Fig. 5
Results of Reactome pathway enrichment, GO analysis, protein–protein interaction, and target protein expressions. A The selected top 25 pathways obtained from Reactome pathway enrichment using DAVID are shown. B Network of drug “targets-pathways” was constructed. The orange square represents a target, and the green diamond represents a pathway. C The results of GO enrichment analysis of the potential targets are shown. D The protein–protein interaction network of the potential targets was generated using STRING. E The differential expressions of DHFR and TYMS in normal and NSCLC tissues obtained from GEPIA database are shown
Table 1 Top 25 representative pathways obtained from DAVIDTo better visualize and quantify the cellular functions of those targets, the 43 potential targets of diosbulbin C in NSCLC were input into the STRING database to construct their protein–protein interactions (PPI). After processing the data by Cytoscape, 41 nodes and 228 edges were identified in the PPI network (Fig. 5D). Notably, AKT1, DHFR, and TYMS show 33, 5, and 9 edges, respectively. The proteins interacting with the above three proteins had a relatively high score, especially the score of DHFR interacting with TYMS was 0.999 (Supplementary Table S3), suggesting that AKT1, DHFR, and TYMS may be the key targets of diosbulbin C in NSCLC. Gene expression analysis of the target genes using the GEPIA online tool revealed significant overexpression of DHFR and TYMS in lung cancer tissues in comparison to the normal controls (p < 0.05) (Fig. 5E). The expression of MMP2, FGFR1, PDGFRB, SELP, CASP1, GSTM2, ALOX5, KDR, FABP4, MET, MMP1, MMP12, MMP13, and MMP9 between lung cancer and normal tissues were also statistically different as analyzed using the GEPIA online tool (Supplementary Figure S2). They may also serve as active targets of diosbulbin C in NSCLC, which need further evaluation.
Molecular docking analysis suggests the good binding affinity of diosbulbin C to AKT, DHFR, and TYMS proteinsGiven the above analysis, three proteins (AKT, DHFR, and TYMS) and diosbulbin C were subsequently subjected to molecular docking analysis using Discovery Studio 2019 (DS). The PBD number of the receptor protein is AKT: 4gv1, DHFR: 1kmv, and TYMS: 3gh0, respectively. The result of molecular docking shows high binding affinities of diosbulbin C to AKT protein by the sites of Val164, Asp292, Glu234, Met281, Lys158, Gly159, Gly162, Thr160, and Phe161 with the -CDOCKER_Interaction_Energy of 49.1404 kcal/mole (Fig. 6A, Supplementary Table S4). Besides, diosbulbin C also shows a high binding affinity to DHFR and TYMS with the -CDOCKER_Interaction_Energy of 47.0942 and 72.9033 kcal/mole, respectively (Fig. 6B and C, Supplementary Table S4). Collectively, the molecular docking results revealed a good binding activity of diosbulbin C with AKT, DHFR, and TYMS proteins.
Fig. 6
Molecular docking and ADMET of diosbulbin C. A Diosbulbin C exhibits high binding affinity to AKT1 protein via sites of Val164, Asp292, Glu234, Met281, Lys158, Gly159, Gly162, Thr160 and Phe161 through molecular docking analysis. B Diosbulbin C exhibits high binding affinity to DHFR protein via sites of Gly117, Gly116, Val115, Phe34, Ile60 and Pro61 through molecular docking analysis. C Diosbulbin C exhibits high binding affinity to TYMS protein via sites of Tyr258, Asp49, His256, Asp254, Ser216, Pro12, His196, Arg185 and Lys107 through molecular docking analysis. D ADMET Plot is plotted by ADMET_PSA_2D vs ADMET_AlogP98. The dark blue dots represent the AlogP98 of diosbulbin C. The red and green ellipses represent 95% and 99% confidence intervals of the human intestinal absorption model, respectively, and the rose red and light blue ellipses represent 95% and 99% confidence intervals of the blood–brain barrier permeability (BBB) model, respectively
In the present study, AKT, DHFR and TYMS, three potential targets of diosbulbin C against NSCLC, were chosen to further analyze the stability of binding to diosbulbin C. The RMSD value represents the positional changes of the protein compared with the initial conformation during the simulation process. The results of MD simulation analysis were shown in Supplementary Figure S3. The systems of AKT, DHFR, TYMS and diosbulbin C were constantly fluctuating, suggesting that the systems were unstable and were changing constantly. However, the fluctuations of AKT and DHFR were relatively small compared to that of TYMS, suggesting the relatively stable binding of diosbulbin C to these targets.
Diosbulbin C downregulates the expression/activation of AKT, DHFR, and TYMS in NSCLC cellsTo further evaluate the effects of diosbulbin C on potential targets, we carried out the qRT-PCR and western blotting analysis. Considering the G0/G1 phase cell cycle arrest induced by diosbulbin C, the expression of AKT, p-AKT, TYMS, DHFR, CDK4, CDK6, Cyclin D1, Cyclin E2, and p-RB proteins, which were highly correlated with cell cycle progression, was analyzed using western blotting. As shown in Fig. 7A, the expression of p-AKT, TYMS, DHFR, CDK4, CDK6, Cyclin D1, Cyclin E2, and p-RB was significantly downregulated both in A549 and H1299 cells treated with diosbulbin C as compared to the untreated groups, further proving the effect of diosbulbin C on cell cycle progression in NSCLC. Besides, the expression of DHFR and TYMS was also analyzed using qRT-PCR, which shows significantly decreased expression in cells treated with diosbulbin C compared with the untreated group (Fig. 7B). These results strongly suggest that diosbulbin C may induce G0/G1 phase cell cycle arrest and inhibit NSCLC cell proliferation by downregulating the expression/activation of AKT, DHFR, and TYMS in NSCLC cells.
Fig. 7
The expression of AKT, DHFR, and TYMS in NSCLC cells after the treatment with/without diosbulbin C. A The expressions of AKT, DHFR, TYMS, CDK4, CDK6, Cyclin D1, Cyclin E2, p-RB in A549 and H1299 cells were detected by western blotting after diosbulbin C treatment for 48 h. B The expressions of DHFR and TYMS were detected by qRT-PCR after diosbulbin C treatment for 48 h. Data are shown as mean ± SD, n = 3. ** p < 0.01, ***p < 0.001, ns: no significant difference
Diosbulbin C exhibits satisfactory predicted ADMET propertiesThe prediction of ADMET properties of a drug is an essential method in contemporary drug design and drug screening. ADMET Descriptors and Filter by Lipinski tools which are available with DS, are employed to predict the ADMET properties of diosbulbin C and assess its drug-likeness. Property parameters, such as aqueous solubility, human intestinal absorption, blood–brain barrier (BBB) penetration, and hepatotoxicity, were generated from the analysis. According to the Lipinski Rule of Five (ROF), which is used to estimate drug-likeness, diosbulbin C exhibits satisfactory ADMET properties (Table 2 and Supplementary Figure S4). Firstly, as demonstrated in Fig. 6D, diosbulbin C is within the 99% confidence interval and is considered reliable for predictions. Diosbulbin C has a molecular weight of 362.374, which is suitable for being developed as a drug. Besides, the ALogP of diosbulbin C is predicted to be 0.639, suggesting a good solubility of diosbulbin C. The numbers of H-bond receptors and donors (six H-bond receptors and two H-bond donors in diosbulbin C) are also within the acceptable range, according to the ROF. Moreover, the levels of solubility, intestinal absorption, and BBB penetration are 3, 0, and 3, respectively, which indicates good aqueous solubility and intestinal absorption, however, low BBB penetration of diosbulbin C (Tables 3 and 4). Cytochrome P450s is a vital enzyme system for drug metabolism in liver. According to the analysis, diosbulbin C is not able to inhibit cytochrome P450 2D6, like most of the chemotherapeutic drugs in lung cancer treatments, such as cisplatin (Table 5). Notably, the result shown in Table 6 suggests no hepatotoxicity would be induced by diosbulbin C. Besides, plasma protein binding (PPB) of a drug can not only determine the diffusion or transport of the drug but also influence its action and efficacy. Our result shows a low plasma protein binding rate of diosbulbin C (Table 7). Moreover, diosbulbin C is predicted to be no mutagenicity and has a rat oral LD50 of 1.11 g/kg, further suggesting its suitability for drug development.
Table 2 The Lipinski Rule of Five of diosbulbin C obtained from the Lipinski tools in DSTable 3 Blood brain barrier penetration and human intestinal absorption prediction of diosbulbin C and reference drugsTable 4 Aqueous solubility prediction of diosbulbin C and reference drugsTable 5 Cytochrome P450 2D6 inhibitor prediction of diosbulbin C and reference drugsTable 6 Hepatotoxicity prediction of diosbulbin C and reference drugsTable 7 Plasma protein binding (PPB) rate prediction of diosbulbin C and reference drugs
留言 (0)