
記住我

Direct, orderly communication between cell populations in distant parts of the brain is made possible by long-range projection neurons (LRPN). The LRPN axons create the brain-wide circuits that enables the highly integrated brain functioning that makes perception, memory, consciousness and skilled movement possible (Sherman, 2016). Depending on their specific branching patterns, LRPN axons can establish parallel (point-to-point) or convergent/divergent (point-to-multi-point) connections, in unidirectional or reciprocal patterns. Importantly, the functional impact of the signals carried by LRPN axons on other cell populations depends on the precise target distribution and the number of their synapses, which is highly diverse and specific for each LRPN type (Clascá, 2022).
Thalamic projection neurons are a key and well-characterized group of LRPN situated deep within the brain. Some of them act as the gateway for sensory information to the cerebral cortex. These neurons cluster together forming the so-called first order relay nuclei of the thalamus, and their axons innervate the ipsilateral cerebral cortex in a focal orderly fashion. One of these nuclei is the ventral posteromedial nucleus of the thalamus (VPM). The VPM projection neurons receive monosynaptic inputs from the trigeminal complex of the brainstem carrying mechano- and nociceptive signals originating in the face and mouth, and relay them to the somatosensory areas of the cortex. In many rodents, this system includes a specialized subsystem that enables these animals to use their motile mystacial vibrissae as high-resolution haptic tactile probes (Bosman et al., 2011).
A number of studies have revealed interesting properties of VPM LRPN neurons and of other thalamic nuclei (Waite, 1973; Saporta and Kruger, 1977; Ito, 1988; Sugitani et al., 1990). The first observation is that these neurons are topographically organized with respect to both their soma location and their axonal terminals in the barrel cortex. The second observation is the existence of region-specific heterogeneous projections from a single nucleus (Han et al., 2018; Muñoz-Castañeda et al., 2021), which suggests the organization of thalamic projections into higher order connectivity motifs (Clascá et al., 2012). These findings indicate that morphological diversity can be found even in a first-order nucleus, which has traditionally been considered to be very homogeneous in terms of the morphology of its neurons.
The concept of motifs plays a prominent role in neuroscience. To provide proper context for the more mesoscopic motifs introduced here, we discuss these prior uses of motifs. In a random network, the directional connections from one neuron to another neuron are made with a certain probability p. The probability for an arbitrary pair to have a single connection is p. The probability for a reciprocal connection is p2. However, when in a network this probability is significantly higher than p2 than we say there are connectivity motifs. This usage (Song et al., 2005) is derived from earlier work in genetic networks (Milo et al., 2002) and has since been applied in many other works (Perin et al., 2011; Vasquez et al., 2013). Another related concept is circuit motifs, which applies to networks comprised of multiple neuron types, such as pyramidal cells, and multiple inhibitory neurons, such as parvalbumin (PV), somatostatin (SOM) and vasoactive intestinal peptide (VIP) expressing neurons and where there is a pattern of connectivity, such as that SOM projects to VIP and PV, but PV does not project to SOM (Pfeffer et al., 2013). Circuit motifs can be related to computations they perform (Womelsdorf et al., 2014) and oscillation frequencies they index (Ter Wal and Tiesinga, 2021). This is in contrast with the projection pattern motifs characterized here, which are related to the presence of higher order structure in projection patterns from individual neurons within a source brain area to multiple target brain areas (Han et al., 2018). This higher order structure should also be statistically significant, meaning that it deviates from the null hypothesis of exclusive first-order projections from a source to target brain areas. According to the null hypothesis, the probabilities of source neurons targeting two or more distinct target brain areas are statistically independent of each other. Significant deviations from this assumption can be considered as evidence for the higher-order organization of LRPN neurons into motifs of projection patterns that are not random.
However, these qualities have been difficult to analyze in experimental data, despite a tremendous development of connectomics approaches in the past decades (Cazemier et al., 2016). On the one hand, it remains impossible to extract single-cell morphologies from a neural population using classical tracing techniques (Oh et al., 2014; Harris et al., 2019) or diffusion tensor imaging (Calabrese et al., 2015). On the other hand, electron microscopy (Kasthuri et al., 2015) and direct synaptic labeling (Druckmann et al., 2014) do not cover a large enough volume to appropriately quantify the morphology of LRPN neurons. These studies instead focus on local circuits, that is, local intracortical projections.
Fortunately, recent developments in automated tissue-to-volume reconstructions of single neurons based on fluorescence micro-optical sectioning tomography (fMOST) (Li et al., 2010) or light-sheet fluorescence microscopy (Economo et al., 2016), enable to image these morphologies at light-microscopic resolution. Despite this progress, characterizing projection motifs in hundreds of single neurons is still challenging and labor intensive. As a result, the sample sizes, even for the most intensively studied LRPN groups are still small, since thousands of neuronal reconstructions are needed to reach a nucleus-wide coverage similar to the mesoscale population experiments by Oh et al. (2014) and Harris et al. (2019). An additional challenge is that topographic maps in relation to high-order connectivity patterns of the brain have not yet been quantitatively characterized, which makes it hard to generalize observations from related data and refine them into mathematically rigorous approaches (Battiston et al., 2020; Bick et al., 2022).
While the existing large-scale repositories have made available large amounts of single-neuron axonal reconstructions (Han et al., 2018; Winnubst et al., 2019; Muñoz-Castañeda et al., 2021; Peng et al., 2021; Gao et al., 2022), they nevertheless have failed to live-up to expectation because each of them were used in isolation. Therefore, data integration from multiple repositories into a collaborative infrastructure, registered into a common reference space and combined with cutting edge neuroinformatics tools for statistical analysis, is necessary for providing context and insight in the morphological distribution and structural organization of thalamic nuclei.
Moreover, to characterize the projection diversity of a neuronal population, we need to go beyond comparing different neurons solely on the basis of their soma position and projection targets. If two neurons with a different soma position in the same source nucleus differ in the targeted areas, there is a number of possible assumptions that can account for that difference. One assumption is that the source region is heterogeneous, with these two neurons belonging to different cell-types, which have different morphologies. The second assumption is that they belong to the same cell-type and that the differences in their projections are due to the position they occupy along the topographical axis within that source region. The last assumption is that the difference is a by-product of the anatomical parcellation scheme, such as the Allen Reference Atlas (ARA) or the Paxinos & Franklin (PF) atlas (Paxinos and Franklin, 2019), which implies that in a different parcellation scheme the two neurons would be classified as targeting the same area. For the first and second assumptions, topographical and morphological analyses will have to be carried out, respectively. For the third assumption, neurons would have to be compared under different parcellation schemes, with the correct one decided by the expert.
For a morphological analysis, morphometrical measures (morphometrics) are used to describe different axonal or dendritic morphological properties of neurons and thus serve as useful features for clustering neurons into types with distinct morphological characteristics (Laturnus et al., 2020; Walker et al., 2022). However, classical morphometrics are inadequate to capture the local topology and geometry of axonal branches and terminals. The reason is that morphometric analyses rely on global pre-defined measurements and do not capture pair-wise local variations between neurons, which obscures the estimation of neuronal variability and biases the global morphometrics employed for this purpose (Kanari et al., 2018; Batabyal et al., 2020). Therefore, when estimating morphological diversity, one needs to take into account the entire axonal tree instead of only regions of interest or pre-computed morphometrics.
To address the aforementioned issues, we have developed a workflow for translating ~300 VPM axonal morphologies into projection statistics with a twofold aim. We analyze the topographical correlations between the VPM and various somatosensory areas and identify distinct subpopulations inside VPM that differ in their connection motifs across the somatosensory areas. The projection statistics are measured according to reconstructed axonal morphologies of long-range projection neurons obtained from two publicly available high-throughput databases, namely MouseLight (Winnubst et al., 2019) and Braintell (Peng et al., 2021). We integrate data from both datasets to deal with the limited sample size of the individual datasets of reconstructed neurons, since together they comprise the most extensive publicly available collection of reconstructed neurons from the thalamus to date.
Moreover, we further characterize the morphological diversity of VPM neurons beyond their projection patterns and in relation to their topographical organization using a robust point-cloud alignment and registration approach called Coherent Point Drift (CPD) (Myronenko and Song, 2010). CPD can compute in an unbiased fashion the morphological distance of two different neurons based on their axonal morphology and anatomical coordinates that goes beyond a given anatomical parcellation. We use CPD to estimate the morphological distance between all possible neuronal pairs, which in turn provides us a distance criterion for defining gradients or clusters that characterize the morphological diversity of VPM. To assess the robustness of CPD-based morphological clusters or types, we overlay them with the corresponding projection types and align them with their topographical orientation along the dorsal-ventral axis. This allows us to analyze the morphologies simultaneously in relation to their topographical, morphological and projection properties. Finally, we implement a number of 2D cortical flatmap and VPM spatial plots for visually inspecting the newly characterized and diversified morphologies in the spatial context of the cortical surface and the VPM nucleus. This analysis could be in principle extended to other types of thalamic projection cells which are structurally more diverse, to help a comprehensive and objective delineation of projection neuron types.
2 Materials and methods 2.1 Data retrieval and neurons reunited portalOver the past 5 years, five repositories of fully traced neurons have been released: the “MouseLight” database (1,500 neurons, Winnubst et al., 2019), the “Braintell” database (1,700 neurons, Peng et al., 2021), the “prefrontal” database (6,300 neurons, Gao et al., 2022) and the smaller collections of visual cortex neurons (46 neurons, Han et al., 2018) and primary motor cortex neurons (38 neurons, Muñoz-Castañeda et al., 2021). These five datasets together comprise currently the most extensive collection of single-neuron long-range projection data, bridging the gap between local microcircuits and brain-wide axonal projections.
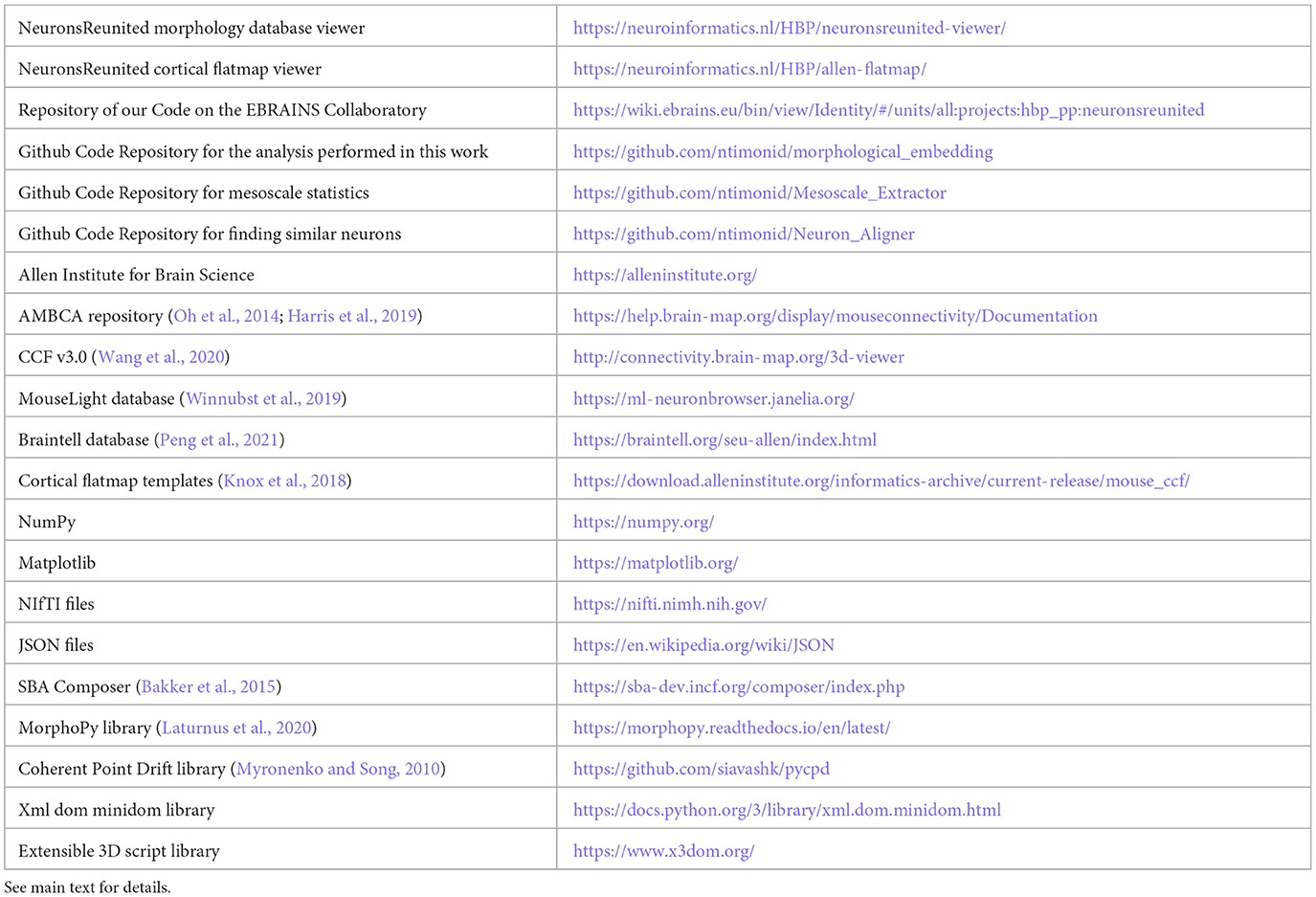
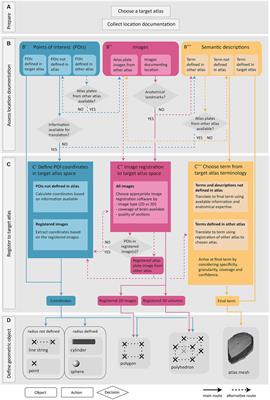
We thus decided to take the opportunity and facilitate access to the community with the Neurons Reunited Portal: a unified access portal to these resources (see Table 1). The website can be used to browse all neurons, and it has a REST application programming interface (API) to pre-select a set of neurons for visualization (see Figures 1A, B). Neurons Reunited Portal is similar to Neuromorpho (Ascoli et al., 2007) in terms of storing neuronal morphologies, with the main difference that all morphologies are by default registered to the common coordinate framework (CCF) v3.0, which allows their direct use in data integration frameworks and spatial statistical analyses such as the ones presented in this work.
Table 1. Hyperlinks for websites, tool descriptions and format descriptions related to our analysis.
Figure 1. Illustration of the different parts of the Neurons Reunited Portal. (A) Partial view of the portal webpage indicating all available databases for visualization and analysis. For each database, the following format has been adopted: the first line describes the name of the database, the second line provides a hyperlink to the respective online publication, the third line is a button enabling the visualization of the somata of all related neurons, the fourth line is a search bar that allows users to search for related neurons with a simple click, and the last line provides hyperlinks for related websites, terms of use, raw data and data citation, if available. (B) Inset of the webpage zooming in on the search bars illustrating how a user can search neurons by database and source brain area. In this example, the keyword “VPM” has been entered in the Braintell search bar. As a result, a list of VPM neurons has appeared to help the user with selecting the appropriate subset of neurons. The acronyms used to characterize each neuron are a combination of the acronyms of taxonomical subdivisions to which the source area of the neuron belongs according to ARA, together with the naming convention provided by the home database of the neuron. For instance, the top acronym appearing on the search bar refers to thalamus (TH), sensory-motor cortex related (DORsm), ventral group of the dorsal thalamus (VENT), ventral posterior complex (VP) and ventral posteromedial nucleus (VPM), while the remaining acronym is the name id of the neuron given by Braintell. (C) Visualization of all available morphologies that are hosted in the portal as clickable spheres. The neurons are color-coded based on their respective database: the light blue color corresponds to Braintell, orange corresponds to MouseLight, red corresponds to the visual cortex database, purple corresponds to the prefrontal database and light green corresponds to the motor cortex database. By clicking on a cell body, its respective morphology gets rendered in a similar fashion to panel (C). The brain template is in horizontal view. (D) Visualization of the selected VPM morphologies using the SBA Composer visualization tool. The red and blue colors correspond to dendritic and axonal, segments, respectively. The brain template is in a sagittal view that is slightly rotated around the superior-inferior axis such that the relationship between axons, dendrites and soma is more clearly visible.
Moreover, the registration of morphologies in CCF enables their visualization in the Scalable Brain Atlas (SBA) composer 3D visualization tool (Bakker et al., 2015), which works as a back-end for the portal. SBA composer can simultaneously visualize all morphologies that are selected by a user, overlaid on a 3D mesh of the CCF mouse brain template (Figures 1C, D). In addition, by clicking on the various brain regions that each neuron originates from or traverses, SBA can render the respective region as a semi-transparent mesh, as well as color-code the laminar distribution of the neurons with different colors, which can significantly enhance the identification and characterization of different sub-types of neurons. Beyond the selection and visualization of morphologies, we have connected the portal with offline Python-based script modules to search for neurons with particular properties, such as “originating in the thalamus and mainly targeting the primary visual cortex“ or with similar morphologies as a number of pre-selected neurons. These enhanced search facilities are part of the workflow presented in this work and have been integrated through bi-directional API interfaces that establish a communication between a Jupyter Notebook analyzing the selected neurons and the Neurons Reunited Portal.
From the above datasets, we stored the complete morphological reconstructions from MouseLight, Braintell and the visual and motor cortex databases. For the prefrontal cortex database, we currently store only the neuronal soma positions. The complete prefrontal reconstructions will be available in the next upgrade of the portal. The original file format of all obtained morphologies is swc (Stockley et al., 1993), which is the de facto format for morphological reconstructions. To store the morphologies in the portal we converted their respective swc files to JavaScript Object Notation (JSON) file formats. The reason for file conversion is because JSON provides a greater degree of flexibility in using Python-related libraries for the analysis described in this work (see Supplementary Section 1.1 for more details regarding the data analyzed in this work).
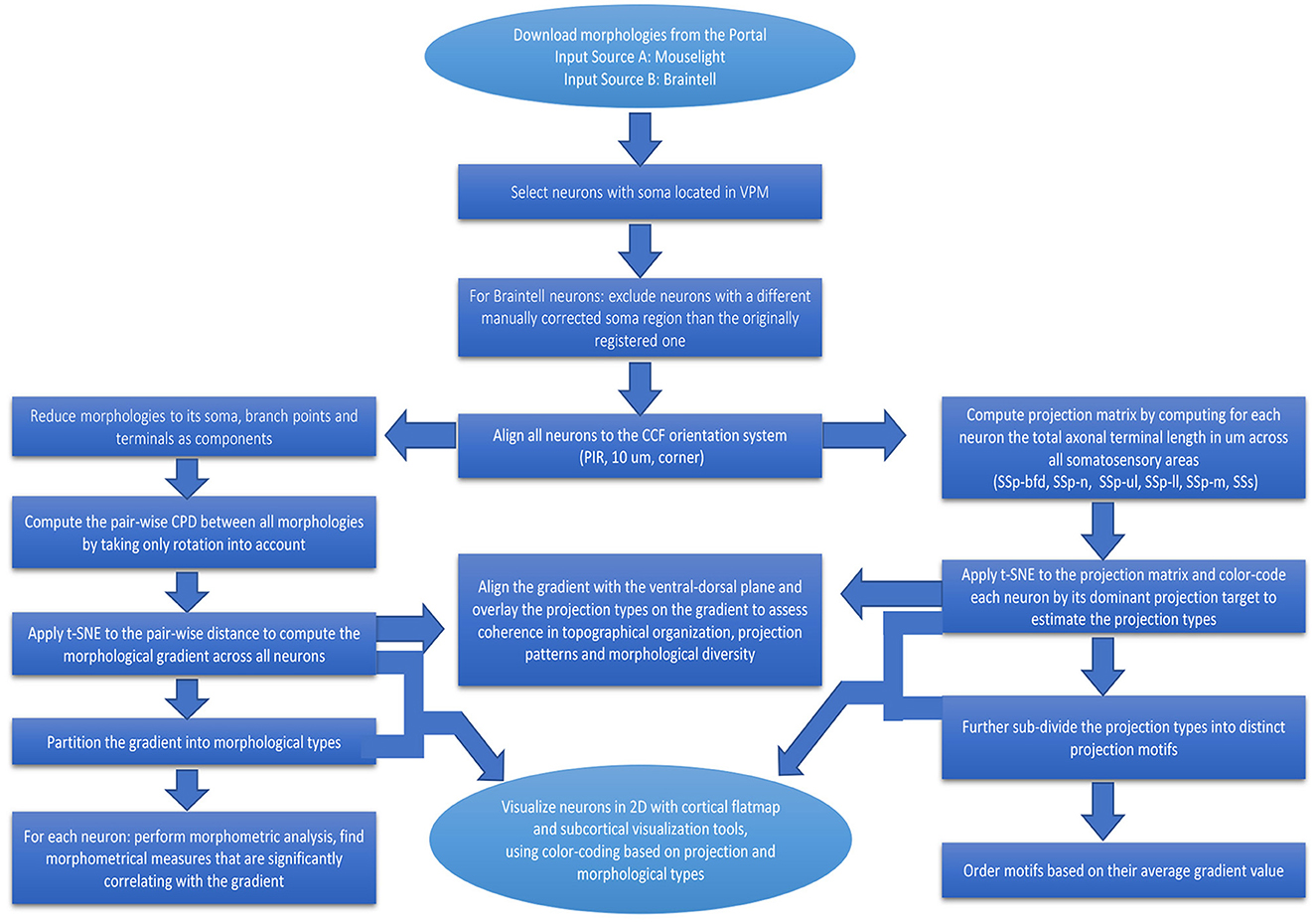
In the workflow described in this study, we analyzed data from two sources of reconstructed neurons from the large online repositories of MouseLight and Braintell. We did not include data from the other databases since they do not contain VPM neurons, which are the focus of this study. A major reason for considering VPM instead of other thalamic nuclei is that it is comprised of a well-studied group of LRPN neurons. Another reason is that it is the most densely sampled nucleus in the thalamus and the second most densely sampled area across the entire mouse brain, when considering all neuronal reconstructions from the MouseLight and Braintell databases together: the first area is the secondary motor cortex (MOs) with 352 neurons and the third area is the caudoputamen (CP) with 256 neurons. In the following sections, we describe different parts of the analysis we conducted to better comprehend the VPM neuronal diversity (see Figure 2 for a schematic description of the various steps comprising the workflow).
Figure 2. Flowchart providing a step-by-step description of our workflow for the analysis of LRPN neurons. Boxes correspond to the various steps comprising the workflow, from pre-processing to analysis and visualization of the data, and arrows represent the directionality of performed actions.
2.2 Data pre-processingWe retrieved data by using an API to download reconstructed neuronal morphologies from the Neurons Reunited Portal, from which 27 were from MouseLight and 256 were from Braintell. The morphologies were downloaded in the JSON file format, for reasons described in Section 2.1. Neurons with a soma position not located in the VPM were filtered out prior to the analysis.
When selecting neurons from the Braintell database, we first had to use a strict selection criterion regarding the registration procedure. For each Braintell neuron, manual corrections were made by Peng et al. (2021) to provide its correct soma area, in order to mitigate errors caused by their fully automated reconstruction pipeline (see Supplementary Section 1.1). However, the corrected 3D coordinates of the soma of each neuron were not released after correction, which would have allowed users to fully utilize the neuron in analyses such the one described here. Therefore, we excluded 26 Braintell morphologies with a manually corrected soma region that did not match the original one. We considered these cases to be potentially misregistered or not originating in the VPM. We thus retrieved 257 morphologies in total for analysis, after applying this selection criterion for all VPM Braintell neurons and after obtaining all VPM MouseLight neurons.
Each morphology was represented as a list of two arrays in which the first corresponds to the anatomical coordinate of each point in CCF and the second contains relevant information about each point, such as its identity (soma, dendrite, axon), its index in the first list and the index of its parent in the list. The parent of a point corresponds to its direct ancestor in the morphological tree, which could be either an axonal branch, dendritic branch or the soma. We label the first array as the “point” array, since its elements are points in 3D space, and the second array as the “line” array, since its elements are lines that connect the points from the first array.
Prior to analysis, we had to ensure for all neurons that the orientation of the coordinate system was the same as for CCF (anterior-posterior, superior-inferior, left-right with origin at the anterior-superior-left corner), by means of an affine transformation. We represented all morphology coordinates at a 10 μm resolution, which is the highest available resolution for the ARA template. First, Braintell neurons are already oriented at PIR (anterior-posterior, superior-inferior, left-right) at 1 μm resolution, with a small fraction having 25 μm resolution. We thus re-scaled the anatomical coordinates of these two Braintell groups by a factor of 10 and 0.4, respectively. Second, MouseLight neurons are oriented at LIP (right-left, superior-inferior, anterior-posterior, corner origin) at 1 μm resolution. For the anatomical coordinates of each neuron, we switched the first and the third coordinate, reflected the third coordinate and re-scaled all coordinates by a factor of 10.
Following the transformation of all neurons to the same reference space, we could now represent each reference neuron as a 3D point cloud. In addition, the parent-child relationship between points allowed us to treat the point-cloud as a morphological tree and hence perform a number of morphometric analyses to it, such as estimating for each axonal segment its path length, radial distance from the soma, branching order, angles, width and height, as well as the number of terminals, thickness and total volume or surface covered.
To alleviate the cost of morphological analysis, we reduced each morphology to its topological minor, which means that it was only represented by its soma, axonal branches and terminals. To identify the axonal terminals, we found axonal points that had no children points. We then extracted the branch of a given terminal by backtracking from the terminal point its ancestor points until a branching point was reached. Therefore, we comprised a list of the axonal terminal branches of each neuron and we integrated them with their branching nodes to preserve the morphological tree structure. The topological minor form allowed us to significantly reduce the computation time of morphological analyses, since it had an on average 30-fold (15,872–411) decrease in number of points used to represent each morphology.
While the above pre-processing steps treated the neurons as morphological trees, we had to create a representation that allowed us to quantify the projection patterns of neurons at the meso-scale level of anatomically distinct subdivisions of the somatosensory cortices, in order to distinguish types of neurons with distinct projection patterns. We therefore defined for each neuron its somatosensory projection targets by estimating the total axonal terminal branch length in μm for each sub-region in the somatosensory cortex: primary somatosensory cortex whiskers/barrel-cortex (SSp-bfd), nose (SSp-n), mouth (SSp-m), upper limb (SSp-ul), lower limb (SSp-ll) and supplemental somatosensory area (SSs). That allowed us to translate morphologies into a connectivity matrix that could be used to define neuronal projection types (see Supplementary Section 1.4 for more details).
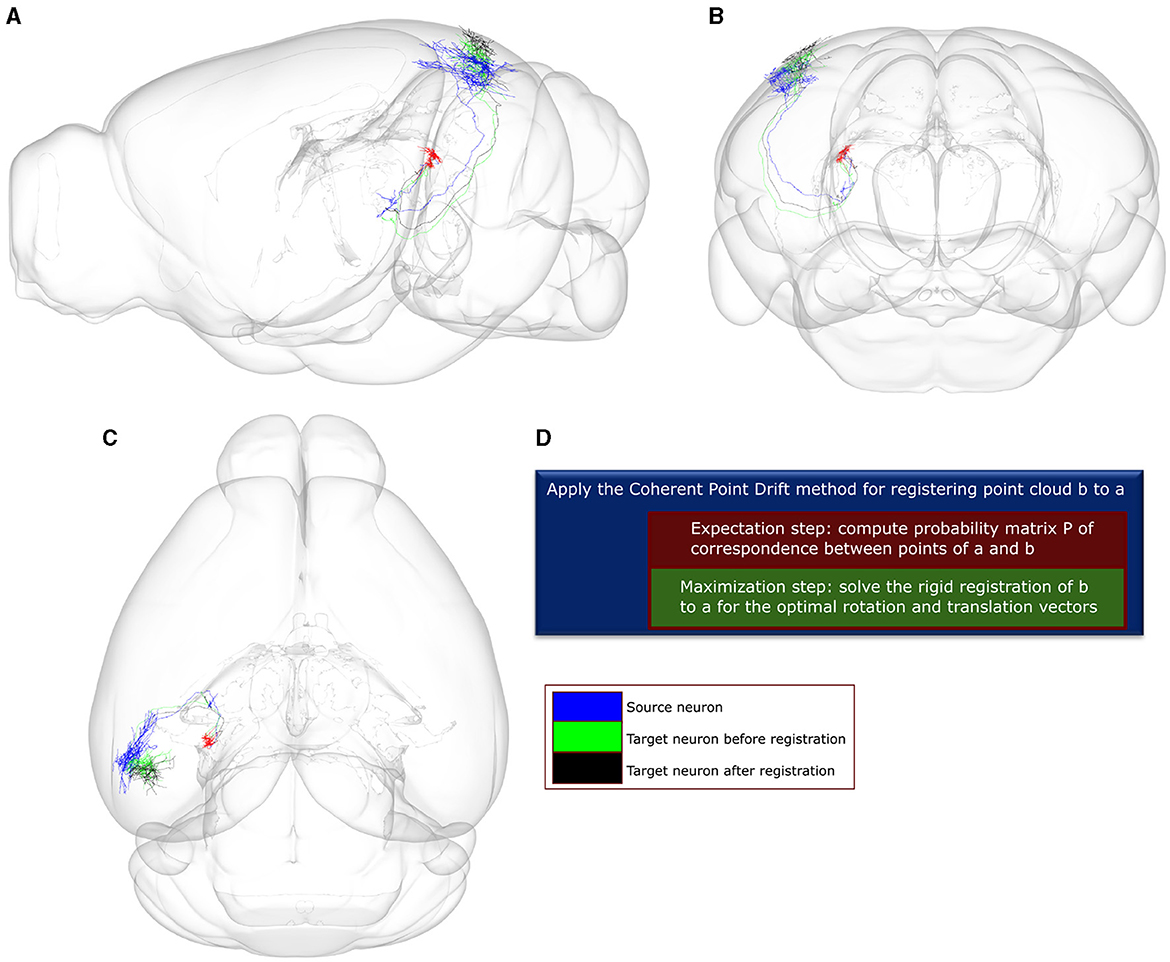
2.3 Searching for similar neurons using the Coherent Point DriftWe then applied the Coherent Point Drift (CPD) method (Myronenko and Song, 2010) to compare all neuronal pairs through a two-step registration process. Given a source and target neuron, CPD finds the missing correspondences between points of the two neurons by minimizing the negative log-likelihood that the point cloud of the source neuron was sampled from the distribution of the target neuron, as modeled by a Gaussian Mixture Model. Subsequently, CPD applies a rigid transformation of the target point-cloud to the source one (Figure 3). The algorithm converges when one of two criteria is met: the algorithm reaches an upper bound of 60 iterations, or the difference between consecutive log-likelihood values falls below the tolerance of 0.001. The values for both criteria were selected to be as strict as possible while achieving convergence under 20 s given an average size of 400 points comprising of the point-clouds used as pairs for the CPD registration. Following convergence, we selected as a neuronal match to each source neuron the target neuron that minimized across all pairs the mean square error (MSE) of registration to the source.
Figure 3. A visualization of the point-to-point correspondence and rigid registration procedure using the Coherent Point Drift (CPD) algorithm, with a use-case involving two example neuronal morphologies a and b that originate in the posterior nucleus of the thalamus (PO). In this use-case, the CPD algorithm is used to find the corresponding points between a and b and subsequently register b to a using rotation and translation. The error for registering b to a is quantified by the mean square error (MSE), which is estimated between each point in b and its corresponding point in a. After 0 and 60 iterations, the MSE is 4.4e+04 and 1.66e+04, respectively. (A–C) Sagittal, coronal and horizontal views of an anatomical mouse brain template based on CCF, to which both neurons have been registered and are spatially overlaid. Green and black colors: axonal branches of neuron b after 0 and 60 rounds of iteration. Blue color: axonal branches of neuron a. Red color: soma locations of both neurons. (D) Description of the CPD algorithm. The high throughput axonal reconstructions have been retrieved from the Braintell repository (Peng et al., 2021). The visualizations were made using the SBA Composer 3D visualization tool (Bakker et al., 2015).
2.4 General methodology for identifying sub-types of neuronsWe have produced a statistical description of the projection motifs of a sensory first-order thalamic nucleus, namely the ventral posteromedial nucleus (VPM) of the thalamus. We characterized neurons in terms of their dominant projection target (Section 2.4.1), and subdivided these groups based on their pattern of projections to secondary areas (Section 2.4.2) as well as on the similarity of the morphology (Section 2.4.3).
2.4.1 Distinguishing projection typesPrior to delineating neurons based on their projection or morphological properties, we applied the t-SNE dimensionality reduction technique (van der Maaten and Hinton, 2008) to create a 2D embedding of the VPM neurons based on their projection patterns across the six somatosensory areas. We thus visualized the data in two dimensions and identified the main sources in axonal variation across the cortex (Figure 4A). Each neuron in its embedded representation was assigned a color that corresponded to the brain area that was its dominant projection target (Figures 4B–E). The dominant projection target of a neuron was defined as the somatosensory area that receives the majority of axonal terminal branches of the neuron compared to the other somatosensory areas. This resulted in a fast, greedy and parameter-free clustering approach which can be applied to quickly pinpoint the dominant projection patterns of the available thalamocortical (TC) morphologies.
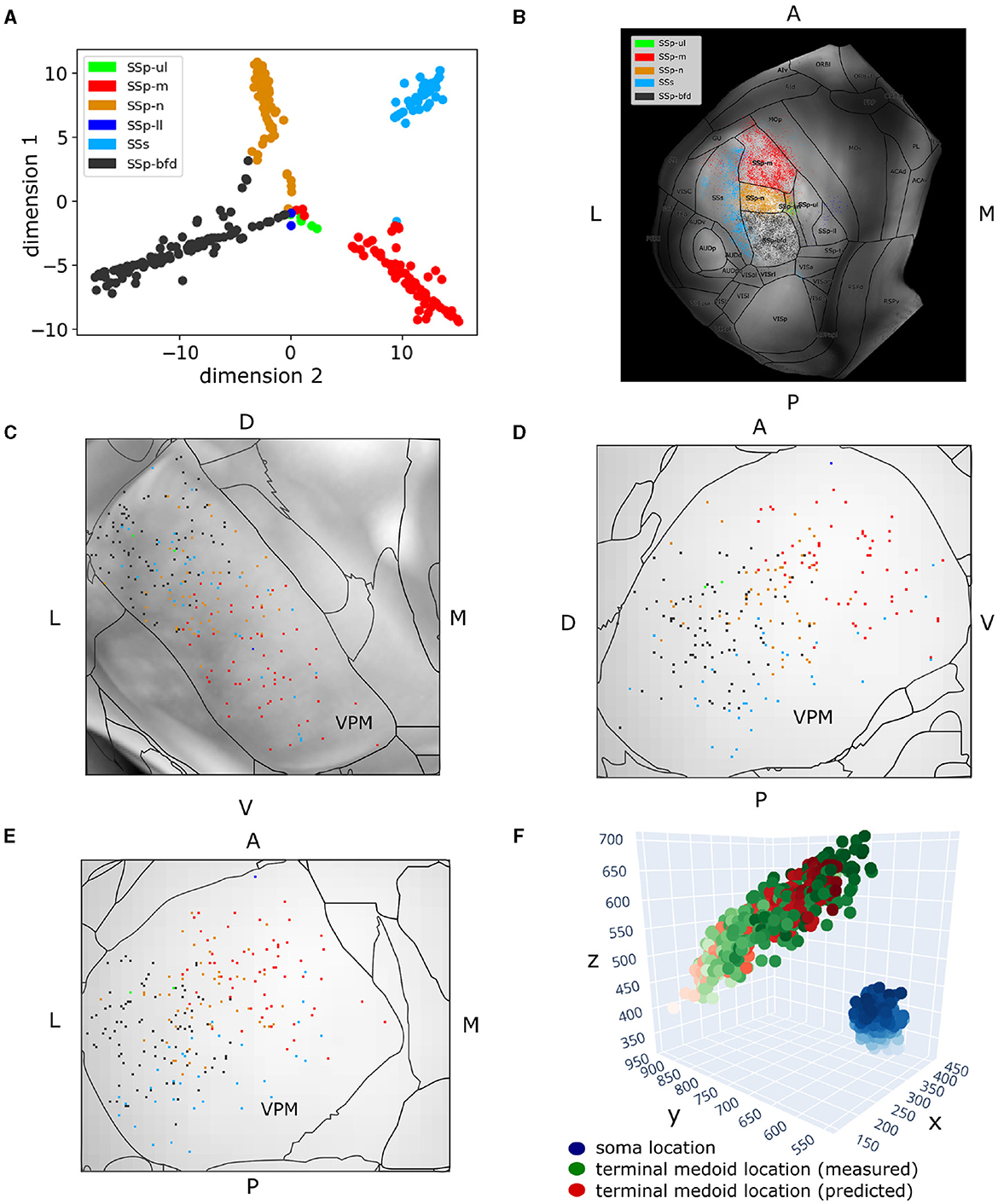
Figure 4. Two hundred and fifty-seven VPM neurons exhibit topographical organization in both their soma and axonal terminal location. (A) Scatter plot illustrating a 2D embedding of the VPM neurons based on their projection patterns across six somatosensory areas (Section 2.4). The projection of each neuron to a given area is defined as the total length of axonal branches that terminate in that area. Each acronym denotes a somatosensory area and the respective color is used to label neurons that have that area as their dominant projection target. The colors are consistent throughout the figure. The x- and y-axes correspond to the projection values of each neuron in the first and second dimension of the embedded space. (B) Dorsal cortical flatmap overlaid with anatomical borders illustrating the axonal termination patterns of the VPM neurons (13.6 × 13.6 mm along the anterior posterior and left-right axes). (C–E) Visualization of neuronal cell bodies in VPM across coronal (C), sagittal (D) and horizontal (E) planes (1.37 × 1.54 × 1.62 mm along the anterior-posterior, superior-inferior and left-right axes, respectively). The letters at the side of each panel denote the plot orientation. (F) 3D scatter plot demonstrating the topographical correlation between the soma position of VPM neurons and their corresponding axonal termination location. Blue color: VPM soma position, with the light-to-dark contrast reflecting the soma position along the ventral-dorsal and medial-lateral axes. Green color: measured axonal terminals, with the light-to-dark contrast reflecting the terminal position along the anterior-posterior and lateral-medial axis. Red color: predicted axonal terminals calculated based on VPM soma position using a least-squares model (see Section 3), with light-to-dark contrast being the same as for the green points. The MSE for reconstructing the measured terminals with this model was 931. The orientation of each axis is shown on its side and follows the RAS orientation: x-axis corresponds to the left-right, y-axis corresponds to the posterior-anterior axis, and z-axis corresponds to the inferior-superior axis of CCF.
To quantify the topographical correlation between the soma positions of VPM neurons and their axonal terminals in the somatosensory cortices, we performed a least-squares regression to fit the soma position of these neurons to the medoids of their axonal terminals. We selected the medoids instead of the centroids to represent the multiple targets that a soma can have, because the medoid measure provides an actual target location that is in the data rather than a location between two or more separated termination domains. By treating the regression coefficient between the two coordinate systems as a rigid transformation matrix, we extracted the corresponding rotation by decomposing the transformation matrix using polar decomposition. We then computed the Euler angles in the x, y, and z axes from the rotation matrix to numerically assess the 3D rotation describing the topographical relationship between the VPM soma positions and their cortical targets (Figure 4F).
Lastly, we assessed the distribution of axonal terminal branch length per layer expressed in μm for each of the four somatosensory areas receiving the highest projection length (barrel field, mouth, nose and supplemental somatosensory area, see Figures 5A–E and Supplementary Figure 1), in order to understand the lamination patterns of each projection type.
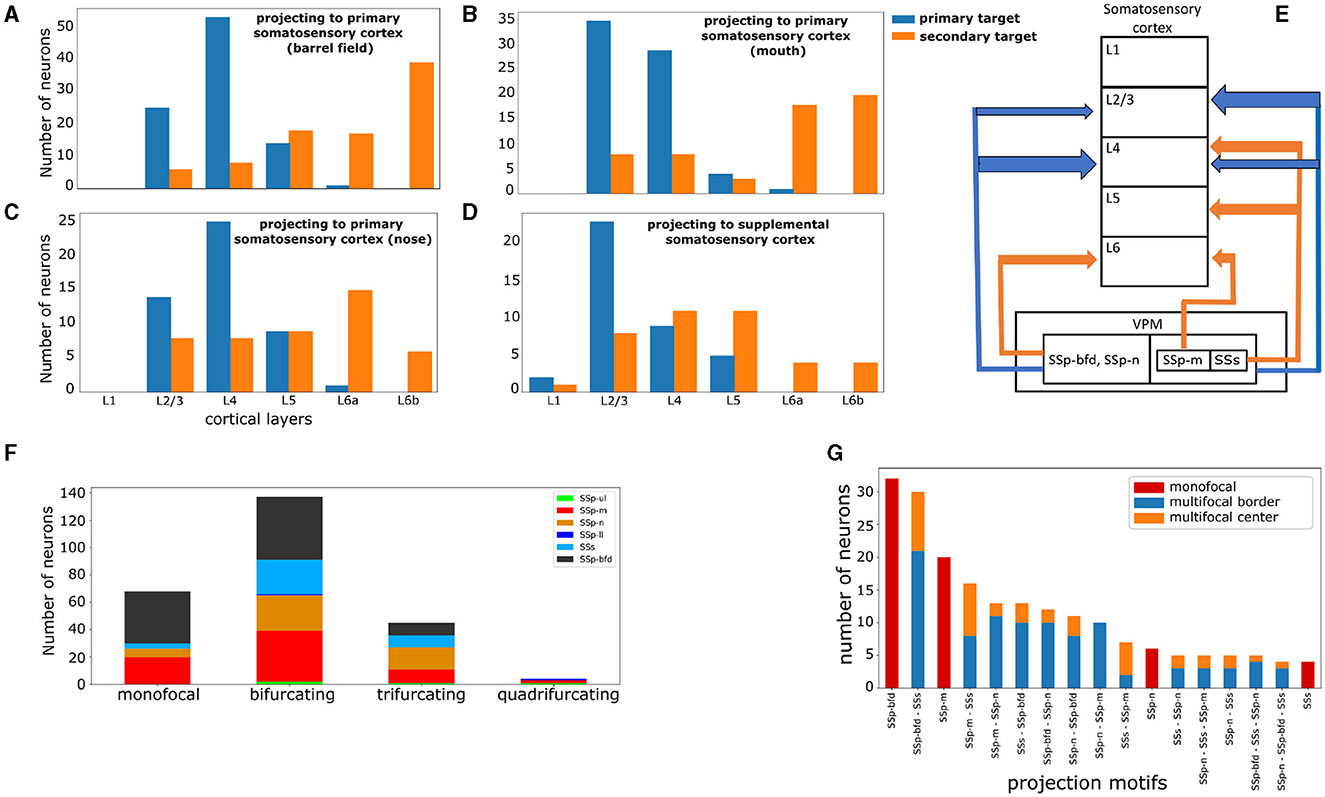
Figure 5. VPM neurons exhibit distinct laminar projection patterns and can be further subdivided by higher order projection motifs. (A–D) Four plots highlighting the cortical layer distribution of the first and second most dominantly targeted area for each VPM neuron, with the first area being either the barrel field (A), mouth (B), nose (C), or supplemental (D) somatosensory area. x-axis: cortical layers. y-axis: number of neurons projecting to a given layer with the highest fraction of terminal length in their first (blue color) and second (orange color) most dominantly targeted area. (E) Schematic illustration that summarizes panels (A–D). The vertical boxes represent cortical layers 1–6 and the nested horizontal boxes represent VPM with its subdivisions based on the projection types that we have delineated in this work. The arrows represent projections to a cortical layer and the arrow thickness represents the relative number of neurons projecting to that layer. The color-coding is the same as in panels (A–D) and it distinguishes primary from secondary projections. The sub-nesting of projection types indicates common projection patterns; SSp-bfd and SSp-n projecting neurons have common laminar projection patterns in both their primary and secondary targets, while SSp-m and SSs projecting neurons share only common primary laminar projection targets. (F) Distribution of neurons based on the total number of jointly targeted brain areas. The motifs exhibit first and higher order connectivity reaching up to three jointly targeted somatosensory areas. x-axis: categories of the different orders of connectivity: monofocal, bifurcating, trifurcating and quadrifurcating correspond to the targeting of one, two, three and four areas, respectively. y-axis: number of neurons belonging to each category. The color-coding is used to distinguish the different projection types and is consistent with Figure 4A, in which the projection types are first introduced. (G) Distribution of the unique projection motifs identified in this work that have been sorted in descending order by the number of neurons participating in each motif and color-coded by the dominant projection target area, with the color-coding as in Figure 4. The areas characterizing each motif are ordered in descending order by the number of axonal terminals received from a single-neuron. The red, blue and orange colors are used to distinguish neurons based on the topography of their projections. Red: monofocal neurons. Blue: multifocal neurons which project in the border between the dominant and the secondary target areas. Orange: multifocal neurons having at least four axonal terminals in the secondary target areas with at least 200 μm distance from the border between these areas and the dominant target area.
2.4.2 Distinguishing projection motifsThe subsequent analysis involved finding projection motifs. A motif was defined by the unique combination of brain areas targeted by the axonal terminals of a given neuron. We identified sub-types based on distinct projection motifs: monofocal neurons projecting to only one area, bifurcating ones projecting to two areas and smaller fractions projecting to more than two areas. For each of the above TC neurons, we obtained the anatomical distribution of their axonal terminals. To filter out weak connections, we estimated which cortical areas received at least five terminal branches for a given neuron. This provided for each neuron the list of areas that it substantially targeted. By grouping neurons based on their common projection lists, we could then highlight the total range of projection motifs that this particular subset of neurons exhibited. From these patterns, we counted the numbers of dedicated inputs to a single-area, as well as bifurcations (i.e., targeting two areas) and trifurcations or higher order motifs (Figure 5F). We found motifs that exhibited first and higher order connectivity reaching up to four jointly targeted somatosensory areas. A preliminary assessment of the statistical significance of VPM projection motifs, which uses the binomial test with Bonferroni correction similarly to how it was achieved in Han et al. (2018), can be seen in Supplementary Figure 1.
We then proceeded to quantifying the distribution of projection motifs in relation to their respective dominant projection types. The areas characterizing each motif were ordered in descending order by the number of received axonal terminals from a single neuron. For instance, motif SSp-bfd - SSs revealed a pattern in which a given neuron most dominantly projected to the somatosensory barrel cortex, while its second most dominant target was the supplemental somatosensory area. We sorted the motifs in descending order by the number of participating neurons to assess the most predominantly targeted areas and we subdivided the motifs in monofocal and multifocal ones to better understand the combinations with which VPM neurons targeted these predominant areas (Figure 5G). A potential issue for multifocal neurons is that their secondary arborizations could be located in the border between the dominant projection area and the secondary one, thus raising concerns about the true nature of such motifs which could be monofocal according to a different anatomical delineation. To address this issue, we further subdivided neurons participating in multifocal motifs in “center” and “border” groups, based on whether they had at least four axonal terminals in the secondary area with a distance >200 μm from the nearest border with the dominant projection area.
2.4.3 Distinguishing morphological gradients and typesWe subsequently applied the Coherent Point Drift method to delineate morphological clusters using the dissimilarity of their axonal trees as a proximity measure (see Section 2.3 for more details and Figure 6A, left for an example). Prior to CPD registration, both neurons were re-centered such that their soma location is located at coordinates 0,0,0. Given two neurons, we computed their dissimilarity by first finding correspondences between the axonal points of both neurons, followed by applying a rigid registration of the former to the latter neuron. We did not allow the registration algorithm to scale the axonal points, which restricted the transformation to only rotating the target neuron. By eliminating the operations of translation and scaling, we focused on finding the optimal rotation required for one morphology to be registered to another. By determining the mean squared distance after convergence of the registration, we ensured the maximum physical proximity between neurons and focused attention solely to the difference in their axonal branching patterns. That allowed us to decouple morphological difference from the neuronal soma locations or their rotation along the inferior-superior axis.
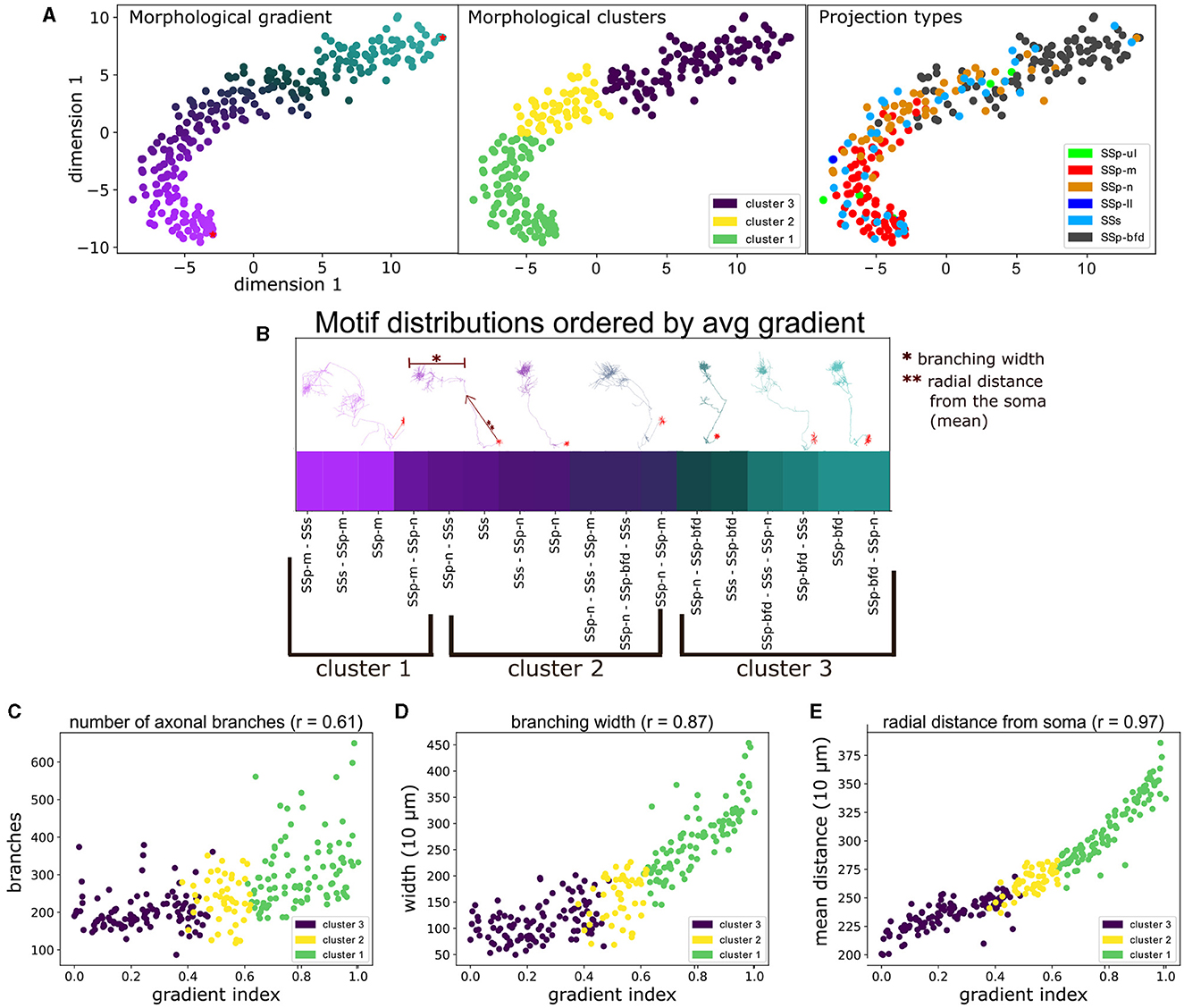
Figure 6. The unbiased characterization of VPM neurons based on their morphological diversity suggests the presence of a gradient that is aligned with their topographical and higher order projection properties, as well as correlated with distinct morphometrical measures. (A) Scatter plot illustrating a 2D embedding of the neuronal morphological diversity (see Section 2.4). The plot has been color-coded three times to highlight the similarity between the three predominant approaches used to distinguish cells in this work, namely by: morphological gradient (A, left), morphologically-defined type (A, middle) and the projection type (A, right) of each neuron. The two red dots at the top and bottom right parts of the gradient in (A, left) indicate the dorsal-most root point and a ventral anchor point, which we used to normalize the gradient scores in the range 0–1. The legend in (A, middle) denotes the color-code of the morphological types, while the legend in (A, right) denotes the color-codes of the projection types, which is described in Figure 4. The x- and y-axes correspond to the morphological values in the first and second dimension of the 2D embedding space. (B) Distribution of the unique projection motifs sorted by the average gradient index of all neurons characterized by each motif (see Section 2.4.2). x-axis corresponds to unique projection motifs and y-axis corresponds to the average gradient index of neurons that are part of a motif. The labels below the x-axis indicate the respective morphological types that characterize the above motifs. The color of each motif corresponds to the color of its respective gradient index as shown in (A). Above the gradient are plotted a number of example neuronal morphologies. Each morphology has the same color as the respective gradient index that sits below its soma position, which highlights the order of the morphology the along the gradient. The arrow next to the second to the right-most morphology and the bar on top of the same morphology are used to exemplify two morphometrics used in this work, the “branching width” and the “radial distance”. The single and double asterisks are used to distinguish the two examples. (C–E) Comparison of the gradient indices of VPM neurons with three significantly correlated morphometrical measures (p < 0.001), namely number of branching points (C), average branching width (D) and average radial distance (E). x-axis corresponds to the gradient indices, y-axis corresponds to values of the respective measure and the r value next to the title of each plot corresponds to the Spearman's r correlation coefficient between gradient and morphometric value: 0.61 for branch points, 0.87 for width and 0.97 for radial distance. The color-coding of all scatter plots is consistent with (A, middle) and indicates the morphological type that each point (neuron) belongs to.
When CPD was performed over all available morphology pairs, we obtained a dissimilarity matrix between all neurons. This matrix could now be used to characterize the morphological diversity between neurons, which was not possible in the space of complete axonal trees containing a variable number of points across different morphologies. The first step was to reduce the dimensionality of the dissimilarity matrix to its two most dominant dimensions in terms of explaining data variance. As described in Supplementary Section 1.4, we used the t-SNE dimensionality reduction technique to the dissimilarity matrix, which estimates a non-linear embedding of the data that respects the topological distance between data points (van der Maaten and Hinton, 2008). This enabled us to visually inspect the morphological diversity in two dimensions with the use of a scatter-plot. We observed that the highest density of the data was along a 1D ridge, hence the second step was to quantify the observed diversity by defining a morphological gradient that can characterize the data, followed by partitioning the gradient into clusters for a simpler morphological characterization.
To define a morphological gradient implied that we could represent each data point on the two dimensional t-SNE embedding using a single number instead of two coordinates, which was inspired by the representation of the data as a curved line in the scatter plot of Figure 6A. Each neuron was assigned a gradient index based on the geodesic distance between its coordinates and the root point coordinates in the 2D embedding space: the root point was assigned to be the point in the upper right end of the t-SNE embedding, which belonged to a barrel-cortex projecting neuron (Figure 6B). The reason for this selection was that barrel cortex projecting neurons were located in the dorsal part of VPM, hence the higher the gradient index the more ventral the neuron was. This provided a positive correlation in Figures 6C–E (see Section 4 for discussion on the topographical analysis). The gradient indices were normalized by the highest index such that they can be represented in the range 0–1.
We then proceeded to check whether there was an alignment between the morphological gradients and the previously identified projection types or motifs. We overlaid the morphological types with the projection types and assessed their overlap (Figure 6A, right). Therefore, we could now pinpoint potential topographical correlations between the soma locations of each cluster and their respective axonal terminals.
We distributed the unique projection motifs identified in this work by sorting them by the average morphological gradient of the neurons belonging to each motif. Hence, we ordered the motifs along their position in the lower embedding morphological space (Figure 6B) and then assessed whether we could place them within a continuum of morphologies defined by the gradient. We then searched for statistically significant correlations between the gradient indices and a number of morphometrical measures, in order to understand whether the morphological differences assessed using our approach were in agreement with pre-computed morphometrics.
To better understand the morphological diversity, we applied hierarchical clustering to the two dimensional embedding to define three clusters or morphological types (Figure 6A, middle). We thus partitioned the gradient into three topographically organized segments that reflect the dorsal, middle and ventral parts in the coronal plane of VPM that most strongly target the barrel, nose and mouth somatosensory cortices, respectively (Peng et al., 2021) (see Section 3.1 for a discussion on VPM topography). We assessed the distribution of the three morphological types across a number of morphometric measures and we computed which morphometrics exhibit significant differences in their distributions across the three types. The motivation was to understand whether the three morphological types exhibited distinct morphological properties and to highlight such properties, which could help us further interpret the morphological types.
2.5 Data visualizationTo proceed in visualizing the results, an appropriate data representation was necessary. Initially we represented the projection or morphological types as two binary 4-dimensional arrays in which the first three dimensions corresponded to the three axes of the 10 μm Allen Reference Atlas: 1,320 voxels in the x-axis representing the anterior-posterior direction, 800 voxels in the y-axis representing the superior-inferior direction and 1,140 voxels in the z-axis representing the left-right directions, with the origin being the anterior-superior-left corner of CCF. The fourth dimension corresponded to the number of neuronal types under analysis. The first 4D array was the source array and encoded the soma positions of all neurons and the second 4D array was the target array and encoded the respective axonal terminal positions. For instance, if the target voxel 0,0,0,0 had the value 1, this means that there existed an axonal terminal from the first neuronal type at the point (0,0,0) in the 10 μm PIR orientation. A value of 1 at the corresponding source voxel would likewise reflect the presence of a soma of the first type at this given coordinate.
Moreover, we implemented a color-coding strategy to simultaneously visualize the different neuronal types. Each type received a unique RGB color as its identification color for any subsequent form of visualization. We could now visually distinguish potential topographically organized patterns found across the different neuronal types, by looking at their color distribution in plots of their source and target regions, prior to proceeding to more numerically rigorous analyses.
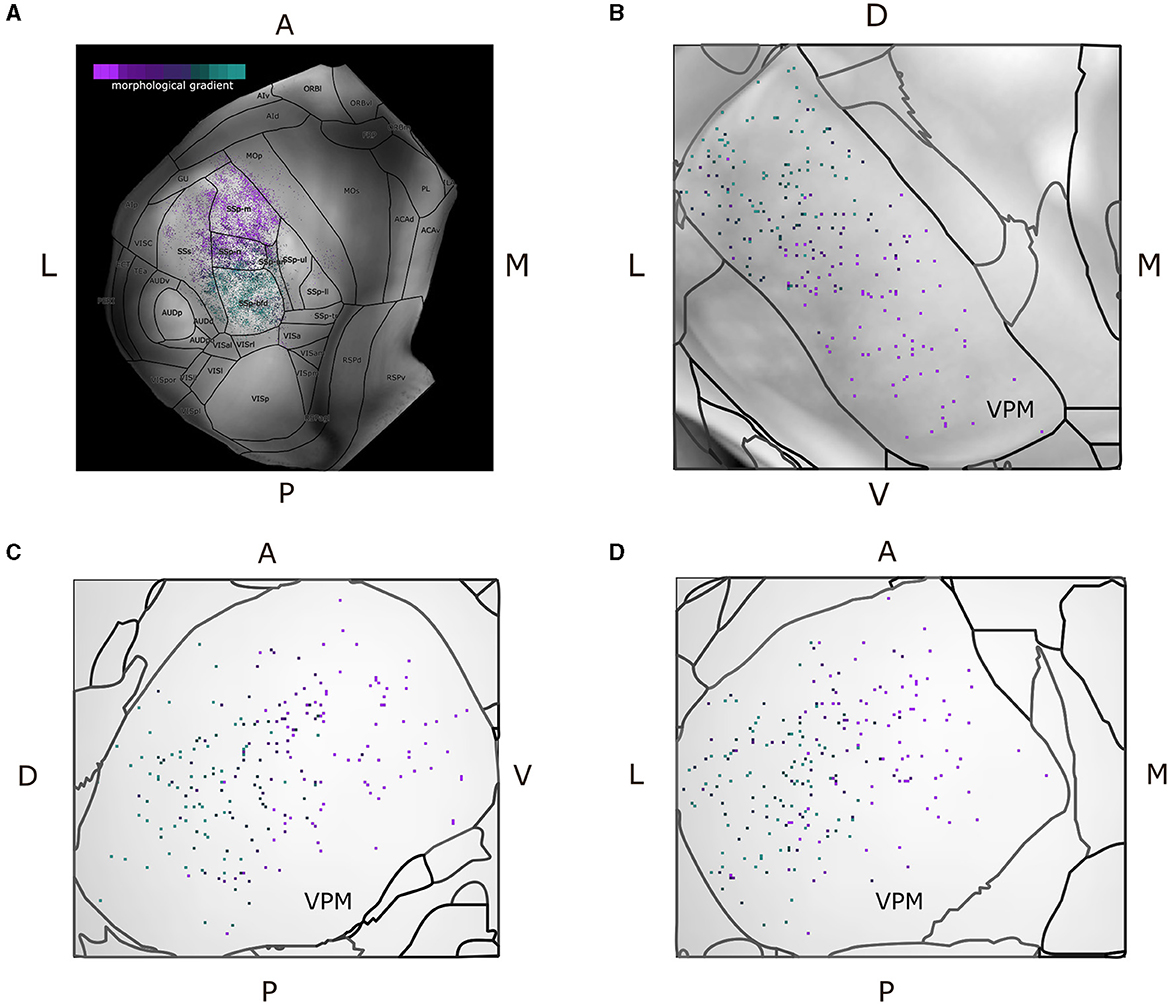
Since we characterized each morphological or projection cluster using different colors, we produced dorsal cortical flatmaps (see Figure 7A) overlaid with anatomical boundaries, as well as 2D projections of VPM (Figures 7B–D, see Supplementary Section 1.2 for the technical implementation details), in order to visually inspect the various clusters. The flatmaps were produced by adapting code from Knox et al. (2018), but we made a number of modifications to improve the anatomical distinction of the projection patterns. In summary, we delineated the anatomical boundaries of ARA using thick black lines, plotted the acronym of each area on its respective center and estimated the flatmap of the STP-based gray matter volumes by taking their average value only across layers 2/3 and 4, such that individual barrels of the barrel cortex become visible. Since the flatmaps are symmetrical and VPM neurons do not have contralateral projections, we created two dorsal flatmaps of only the left hemisphere to reduce memory usage. We therefore mirror the axonal termination patterns of the right hemisphere to the left flatmap one.
Figure 7. Dorsal cortical flatmap and maximum projection plots of VPM verify the topographical distribution of the morphological gradient that characterizes VPM neurons. (A) Dorsal cortical flatmap illustrating the axonal termination patterns of the VPM neurons (13.6 × 13.6 mm along the anterior posterior and left-right axes). The color-coding shown in the top left legend is consistent across panels and is the same used for the morphological gradient index in Figure 6. The correlation between the morphological gradient and the gradient of axonal terminals along the anterior-posterior, lateral-medial axes, as defined by Spearman's r, is −0.8, while the equivalent correlation with the gradient of soma positions along the ventral-dorsal, medial-lateral axes is −0.83 (p < 1e-05). The color-coding strategy used consistently throughout figures (A–D) is explained in the inset of panel (A). (B–D) Visualization of neuronal cell bodies in VPM across coronal (B), sagittal (C), and horizontal (D) planes (1.37 × 1.54 × 1.62 mm along the anterior-posterior, superior-inferior and left-right axes, respectively). The letters at the side of each panel denote the plot orientation.
To represent the VPM soma position distribution in 2D, three maximum projection plots were created for the coronal, sagittal and horizontal planes, respectively. For each plane, we overlaid the maximum projection of three different components along the plane's axis: first, the color-coded locations of the somata, second, the background gray-matter volume from the STP-images of the CCF v3.0 template (Wang et al., 2020) and third, contour lines that delineated the anatomical borders of VPM according to the ARA parcellation (Figure 7C). Lastly, we imported all the morphologies that were of interest directly to the Scalable Brain Atlas (SBA) composer (Bakker et al., 2015) tool, which enabled an easy-to-use 3D rendering of the data within the CCF template. In SBA, we color-coded the morphologies using the same strategy as described above (see Supplementary Section 1.5).
3 ResultsBased on the collection of reconstructed neurons collected here we have identified a topographical correlation between the location of source somata in VPM and their arborization patterns in the somatosensory cortices. We analyzed this topographical correlation and identified different neuronal populations inside VPM that preferentially targeted different combinations of somatosensory areas. In the paragraphs below, we will describe these findings in more detail.
3.1 VPM neurons can be characterized by their topographical organization, layer target specificity and higher order projectionsWhen embedding in two dimensions the total terminal length per neuron across the somatosensory areas, four of the six areas appeared to be dominantly targeted, namely SSp-m, SSp-n, SSp-bfd and SSs (see Section 2.2 for notations). The data created four sharp clusters, with each cluster being almost completely homogeneous in terms of the dominant projection target of its neuron members; the only exceptions are a few neurons located in the middle of Figure 4A having the SSp-ll and SSp-ul as their dominant projection targets. Therefore, we concluded that neurons could be characterized as dominantly projecting to the primary somatosensory areas representing mouth, nose, whiskers or the supplemental somatosensory area.
When overlaying the projection types onto a dorsal cortical flatmap (Figure 4B), they appeared to be topographically organized. This was evident by the minimal presence of projections outside the dominant projection target, which occurred mostly in the borders between areas, with the exception of the SSp-m neurons targeting the lateral part of SSs. The various somatosensory targets (barrel field, mouth, nose, supplemental) were seen to originate in spatially distinct subareas of VPM. Along the dorsal-ventral and lateral-medial planes inside VPM, neurons appeared to target the barrels, nose and mouth, respectively, which indicated a similar topographical organization of the soma distribution (Figures 4C–E). This finding has been extensively described in the rat VPM (Waite, 1973; Saporta and Kruger, 1977; Ito, 1988; Sugitani et al., 1990) and more recently in the mouse VPM (Peng et al., 2021).
To validate this observation numerically, we fitted for all neurons their soma position to the medoids of their axonal terminal points and extracted the Euler angles describing the rotation required for the former to match the latter, as described in Section 2.4.2. The MSE of transforming the soma coordinates to their target medoids at 10 μm resolution was 931, which we considered adequate (see Figure 4F). The corresponding Euler angles in the x, y, and z axes, respectively, were −62.8, −1.5, −126.8. We thus deduced that for the VPM somata to match their target centers, they have to perform a negative rotation around the anterior-posterior and lateral-medial axis, with an almost zero rotation around the dorsal-ventral axis. This finding quantitatively validates the above observation that dorsal-ventral and lateral-medial soma positions in VPM correspond to posterior-anterior and lateral-medial axonal terminal locations in the somatosensory cortex.
Regarding preferential laminar targets, a clear majority of VPM neurons projected to layers 2/3 and 4. In SSp-bfd, SSp-n and SSs, layer 4 was the most dominantly targeted layer with 5,546, 5,022 and 3,487 μm of axonal terminal length, respectively, with layer 2/3 being the second most targeted cortical layer (Supplementary Figures 2A, C, D). In SSp-m, layer 2/3 was the most dominantly targeted layer with 3319 μm of axonal terminal length, with layer 4 being the second most targeted cortical layer (Supplementary Figure 2B). When taking into account the layer to which neurons project their highest fraction of axonal terminals length, we identified layer 4 as the selected layer for the SSp-bfd and SSp-n projection-types (53 and 25 neurons, respectively) and layer 2/3 for the SSp-m and SSs projection-types (35 and 23 neurons, see Figures 5A–D). In addition, when assessing the layer with the highest terminal length for the secondary target of multifocal neurons, we identified layer 6b for SSp-bfd and SSp-m (39 and 20 neurons), layer 6a for SSp-n (15 neurons) and layers 4 and 5 for SSs (11 neurons each, Figure 5E).
Regarding interpretation of these findings, projections to layer 4 are well established as the “core” projections that carry feed-forward input from the lemniscal system to the cortex and have been shown to display driver-like characteristics (Sherman, 2016; Harris et al., 2019; Clascá, 2022). Inputs to layer 2/3, although predominantly modulatory, can also evoke the firing of action potentials in some cells, suggesting that both layers participle in the encoding of active touch through direct signals from the thalamus (Crochet et al., 2011; Viaene et al., 2011). Moreover, neurons projecting to layer 2/3 have a shorter length of arbors than the layer 4 projecting ones, as well as shorter projections to the deeper layers, as has been previously reported in Peng et al. (2021). The functional relevance of these secondary projections to the deeper layers is something that has not been experimentally explored in VPM, which has been traditionally considered to be a sensory relay nucleus with point-to-point connections. However, this will be further elaborated in Section 4. Lastly, the secondary projections to layers 2/3 and 4 by multifocal SSs-projecting neurons can be explained by most of these neurons having SSp-m or SSp-bfd as their second most dominantly targeted area, which predominantly receive input in layer 2/3 or 4, respectively, as described above.
Regarding the broadcasting properties of VPM neurons, 26% of VPM neurons were found to be monofocal, with 53% being bifurcating, while smaller fractions of neurons were tri- or quadrifurcating (17 and 1.5%, respectively, see Figure 5F). SSp-m, SSp-n, SSs receive markedly more multifocal projections relative to their monofocal ones (49 multifocal over 20 monofocal projections for SSp-m, 42 over 6 for SSp-n and 34 over 4 for SSs), while SSp-bfd has a more balanced ratio of multifocal to monofocal projections (55 over 38). The presence of monofocal VPM neurons sending dedicated projections to specific somatosensory areas, with individual barrels being the most predominant target, is well known in literature. For instance, in rats it has been reported that 70% of barrel-cortex projecting VPM neurons target individual barrels and 30% targets multiple barrels, with the latter located in the marginal regions of VPM (Sugitani et al., 1990). This raises the question about the presence and function of neurons from first-order nuclei targeting multiple somatosensory areas instead of exclusively targeting a single area (Clascá et al., 2012). To better understand these bifurcated projections, we put the projection motifs within the context of their dominant projection targets and we overlaid them with the morphological gradients, which we discuss in the following paragraphs.
When observing the entire spectrum of VPM projection motifs, 17 out of 40 motifs in total were substantially represented by a
留言 (0)