
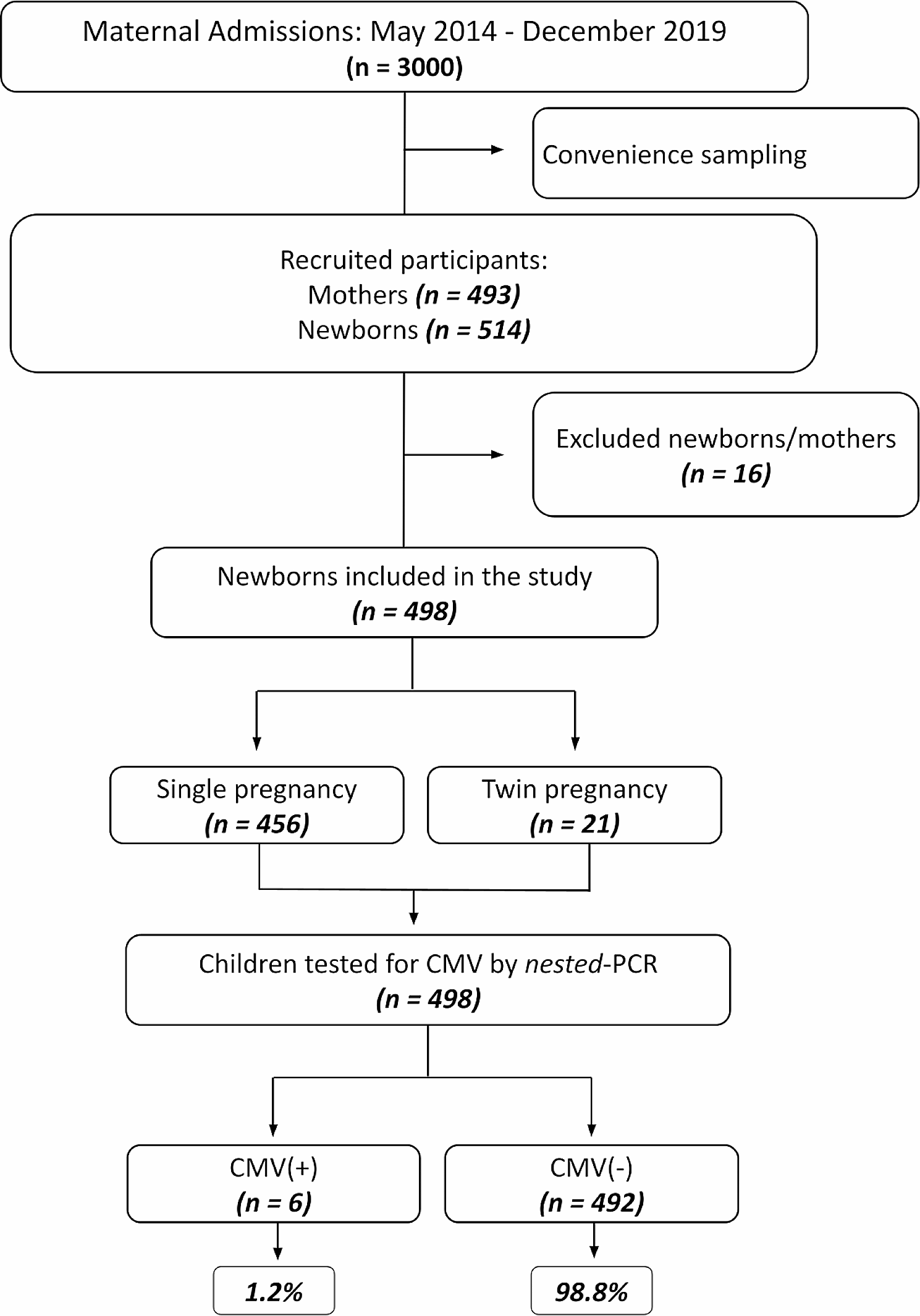
記住我





The conserved octamer motifs are identified and found to be located 70 to 90 nucleotides upstream of the poly(A) tail in coronaviruses from different genera based on the analysis of the available coronavirus sequences in GenBank [9, 11, 22, 24]. The examples of the conserved octamers derived from different genera of coronaviruses are shown in Table S1, although few coronavirus species contain one to three nucleotides that differ from those in the conserved octamer GGAAGAGC.
Based on a previous study [22], MHV-A59 with point mutation and complete deletion of the octamer motif can replicate well in cultured cells. In addition, MHV-A59 with complete deletion of the octamer motif has been demonstrated to be attenuated in mice [22]. However, whether MHV-A59 with complete mutation of the octamer motif (that is, mutation from GGAAGAGC to CCUUCUCG) can also replicate to high titers and whether such a mutation can lead to attenuation in mice remain unknown. Accordingly, MHV-A59 with complete mutation of the octamer motif was selected for the current study. To examine whether the coronavirus with complete mutation in octamer motif had the aforementioned features, wt octamer GGAAGAGC was mutated to CCUUCUCG and thus wild-type (wt) MHV-A59 (MHVwt) and MHV-A59 octamer mutant (MHVoctm) full-length cDNA (Fig. 1A) were constructed. Different multiplicities of infection (MOIs) of MHVoctm and MHVwt ranging from 1 to 0.0001 were used to infect ML cells, and supernatants were collected at different time points of infection. As shown in Fig. 1B-N, it was found that (i) at 8 hpi, with the reduced MOI from 1 to 0.0001, the virus titers for both MHVwt and MHVoctm were significantly decreased (from 3.0 × 106 to 3.9 × 101 for MHVwt and from 3.5 × 105 to 1.4 × 101 for MHVoctm), and the virus titer for MHVoctm was undetectable at an MOI of 0.0001 (Fig. 1B-F); (ii) at 8 hpi, with the reduced MOI from 1 to 0.001, the difference in virus titer between MHVwt and MHVoctm was significant (8.3- to 11.5-fold) (Fig. 1L); (iii) at 16 hpi, with the MOI reduced from 1 to 0.001, the virus titer was slightly decreased for MHVwt (from 3.9 × 107 to 8.1 × 105) and MHVoctm (from 3.7 × 107 to 3.4 × 105) (Fig. 1B-E), and the difference in virus titer between MHVwt and MHVoctm was slightly increased (from 1.0- to 2.4-fold) (Fig. 1M); however, with 0.0001 MOI, the difference was significantly increased (36.9-fold) (Fig. 1F, K and M) ; (iv) at 24 hpi, even with the reduced MOI, the virus titer for both MHVwt and MHVoctm was high and similar (Fig. 1B-F), and the difference was only slightly increased (from 1.1 to 2.4 folds) (Fig. 1N). Therefore, with a reduction in MOI from 1 to 0.0001, the replication efficiency for both MHVwt and MHVoctm was significantly decreased during the earlier stage of infection (especially at 8 hpi). In addition, the difference between MHVwt and MHVoctm in virus titer during the earlier stage of infection (8 and 16 hpi) was increased. Notably, with 0.0001 MOI, the virus titer for MHVoctm was even undetectable at 8 hpi and the difference was significantly increased (36.9-fold) at 16 hpi. However, the virus titer for both MHVwt and MHVoctm was high and similar at the later stage of infection (24 hpi). The results suggest that, with reduced MOI for infection, the difference between MHVwt and MHVoctm in virus titer at the earlier stage of infection (8 and 16 hpi) become evident; however, even with mutated octamer and reduced MOI for infection, MHVoctm still can grow to high titer similar to MHVwt in cultured cells at the later stage of infection, establishing MHVoctm as a potential candidate for the vaccine development.
Fig. 1
Growth kinetics of octamer-mutated coronavirus and wt coronavirus at different multiplicities of infections in cultured cells. (A) The genome structure of wt MHV-59 (MHVwt) and MHV-A59 with octamer mutation (MHVoctm). (B)-(F) Growth kinetics of MHVwt and MHVoctm in ML cells infected with MOIs of 1, 0.1, 0.01, 0.001 or 0.0001. (G)-(K) The folds of virus titer of MHVwt over MHVoctm (wt/octm) at different times postinfection (8,16, and 24 hpi) with MOIs of 1, 0.1, 0.01, 0.001 or 0.0001 derived from Figures (B)-(F). (L)-(N) The folds of virus titer of MHVwt over MHVoctm (wt/octm) with different MOIs at 8 (L), 16 (M) and 24 (N) hpi derived from Figures (B)-(F). The values in (B)-(N) represent the mean ± standard deviation (SD) of three individual experiments. Statistical significance was evaluated using a t test: * P < 0.05, ** P < 0.01. MOI, multiplicity of infection; hpi, hours postinfection; MHVwt or wt, wild-type MHV; MHVoctm or octm, MHV with octamer mutation
The coronavirus with mutated octamer leads to the reduced and lagged synthesis of viral proteinsSince (i) MOI can affect the replication difference between octamer-mutated (MHVoctm) and wild-type (MHVwt) coronavirus during infection (Fig. 1), (ii) the mice may be infected with much lower MOI of virus than cultured cells and (iii) the efficiency of viral proteins synthesis is also a factor affecting the pathogenicity, it is hypothesized that, with the reduced MOI, the decreased translation efficiency of MHVoctm in comparison with that of MHVwt at the earlier stage of infection may suggest its potential to be attenuated in mice, supporting its role as a vaccine candidate. To examine the hypothesis, different MOIs of MHVoctm and MHVwt ranging from 1 to 0.0001 were used to infect ML cells, and cell lysates were collected at different time points of infection. As shown in Fig. 2B, with a higher MOI of 1, in comparison with MHVwt, MHVoctm showed reduced translation efficiency at 8 h postinfection (hpi), but had almost the same translation efficiency at 12, 14, 16 and 24 hpi. With a reduction in MOI from 0.1 to 0.0001, the synthesis of detectable viral proteins was delayed (Fig. 2C-F). In addition, the translation difference between MHVoctm and MHVwt was observed, and the occurrence of the difference was also delayed (Fig. 2C-F). Consequently, the results suggest that, with reduced MOI for infection in cultured cells (Fig. 2B-F), (i) the synthesis of detectable viral proteins gradually lagged after infection and (ii) the difference in translation efficiency between MHVwt and MHVoctm became significant. Since translation occurs prior to replication, the difference in virus titer with different MOI at the earlier stage of infection (Fig. 1) may be at least partly due to the effect of the octamer mutation on translation efficiency. Consequently, the results further suggest that the octamer-mutated MHVoctm has the potential to be attenuated in mice and thus is a potential vaccine candidate.
Fig. 2
The coronavirus with mutated octamer leads to the reduced and lagged synthesis of viral proteins. (A) The genome structure of wt MHV-59 (MHVwt) and MHV-A59 with octamer mutation (MHVoctm). (B)-(F) Detection and comparison of N protein synthesis between MHVwt and MHVoctmin in ML cells infected with different MOIs (1, 0.1, 0.01, 0.001 and 0.0001, B-F) at different times postinfection by Western blotting. The amounts of the N protein were normalized to those of actin. The values in (B)-(F) represent the mean ± standard deviation (SD) of three individual experiments. Statistical significance was evaluated using a t test: * P < 0.05, ** P < 0.01, **** P < 0.0001. MOI, multiplicity of infection; M, mock infection; N, nucleocapsid protein; hpi, hours postinfection; MHVwt or wt, wild-type MHV; MHVoctm or m, MHV with octamer mutation
The coronavirus with mutated octamer is more sensitive to interferon treatment than wt coronavirusIt has been demonstrated that coronavirus-encoded protein can antagonize host innate immunity [32]. Consequently, since MHVoctm with mutated octamer displays less translation efficiency than MHVwt (Fig. 2), it was hypothesized that the reduced translation efficiency for octamer-mutated MHVoctm may have a weaker ability to weather the challenge of innate immunity and thus reduce the replication efficiency, which would also be a potential factor leading to the attenuated phenotype of octamer-mutated MHVoctm in vivo. To compare the capability against the challenge of innate immunity between MHVoctm and MHVwt, ML cells were first treated with different doses of interferon (IFN) β followed by infection with MHVwt or MHVoctm at an MOI of 0.01. As shown in Fig. 3A and summarized in Fig. 3D, without IFNβ treatment, the virus titer of MHVwt was higher than that of MHVoctm at 8 and 16 hpi but was similar to that of MHVoctm at 24 hpi. However, with the treatment of 104 U/ml IFNβ, the difference in virus titer between MHVwt and MHVoctm was increased with time (Fig. 3C and summarized in Fig. 3F). In addition, with increased amounts (from 0, 103 to 104 U/ml) of IFNβ treatment, the difference in virus titer between MHVwt and MHVoctm was also increased at 16 and 24 hpi (Fig. 3H and I). The results together suggest that MHVoctm is more sensitive to IFNβ treatment than MHVwt.
Fig. 3
Octamer-mutated MHVoctm is more sensitive to interferon treatment than wt coronavirus MHVwt. (A)-(C) The growth kinetics of wt MHV-59 (MHVwt) and MHV-A59 with octamer mutation (MHVoctm) at 0.01 MOI with 0 (A), 103(B) or 104(C) U/ml of IFNβ. (D)-(F) The folds of virus titer of MHVwt over MHVoctm (wt/octm) at different times postinfection (8,16, and 24 hpi) with different amounts of IFNβ (0 U/ml (D), 103 U/ml (E) and 104 U/ml (F)) derived from Figures (A)-(C). (G)-(I) The folds of virus titer of MHVwt over MHVoctm (wt/octm) with different amounts of IFNβ (0, 103 and 104 U/ml) at 8 (G), 16 (H) and 24 (I) hpi derived from Figures (A)-(C). wt, wild-type MHV-A59; octm, MHV-A59 with octamer mutation; hpi, hours postinfection. Statistical significance was evaluated using a t test: ** P < 0.01, *** P < 0.001, **** P < 0.0001
Further examination revealed that, with IFNβ treatment, the capability of inhibiting the IFNβ signaling pathway for MHVwt was overall stronger than that for MHVoctm because the mRNA levels of IFNβ (Fig. 4A-C), 2′,5′-oligoadenylate synthetase (OAS) (Fig. 4D-F) and interferon-stimulated gene 15 (ISG15) (Fig. 4G-I) in IFNβ-treated and MHVwt-infected ML cells were overall lower than those in IFNβ-treated and MHVoctm-infected ML cells. Taken together, the results suggest that MHVoctm is more sensitive to IFNβ treatment than MHVwt. Since coronavirus-encoded proteins can antagonize innate immunity [32], the weaker capability of MHVoctm to inhibit innate immunity may also be a potential factor leading to the attenuated phenotype of octamer-mutated MHVoctm in mice.
Fig. 4
The capability of inhibiting the IFNβ signaling pathway by MHVwt and MHVoctm. (A)-(I) Relative amounts of IFNβ (A)-(C), OAS (D)-(F) and ISG15 (G)-(I) mRNA synthesis in ML cells treated with different amounts of IFNβ followed by mock infection or infection with MHVwt or MHVoctm (0.01 MOI) at different times postinfection. “Mock” on the y-axis indicates the amount of mRNA detected from IFNβ-treated and mock-infected cells. The fold change/mock on the y-axis is presented as relative units of mRNA compared to the amount of mRNA in IFNβ-treated and mock-infected cells (the amount of mRNA in IFNβ-treated and mock-infected cells = 1). The values in (A)-(I) represent the mean ± standard deviation (SD) of three individual experiments. Statistical significance was evaluated using a t test: * P < 0.05, ** P < 0.01, *** P < 0.001, **** P < 0.0001. hpi, hours postinfection; IFN, interferon; OAS, 2 ′,5′-oligoadenylate synthetase; ISG15, interferon-stimulated gene 15; MHVwt or wt, wild-type MHV; MHVoctm or octm, MHV with octamer mutation
Coronavirus with octamer mutation is attenuated in miceBased on the results shown in Figs. 1 and 2, and 3, the octamer-mutated MHVoctm has the potential to be a vaccine candidate. Therefore, to verify whether the octamer-mutated MHVoctm can be attenuated in mice, 3-week-old male BALB/c mice were intraperitoneally inoculated with Dulbecco’s Modified Eagle Medium (DMEM), 106 pfu of MHVwt or MHVoctm (Fig. 5A). Substantial weight loss, clinical signs and death were not observed in mice inoculated with MHVoctm but MHVwt (Fig. 5B-D). The levels of viral titers, proteins and RNA were also lower in MHVoctm-infected mice than in MHVwt-infected mice (Fig. 5F-H). In addition, the results shown in Fig. 5G demonstrated that MHVoctm could express viral N proteins, although the amounts of the proteins expressed by MHVoctm were much lower than those expressed by MHVwt. Thus, the results suggest that MHVoctm can replicate in mice. Histopathological examination revealed coagulation necrosis and vacuolated hepatocytes (ballooning degeneration) in the livers of mice inoculated with MHVwt, but no histopathology changes were observed in mice inoculated with MHVoctm (Fig. 5I). Together with the results obtained from cell cultures shown in Figs. 1, 2 and 3, the findings indicate that the dramatically decreased gene expression of MHVoctm in mice may be at least partly attributable to the mutated octamer and the reduced MOI, which led to decreased gene expression and thus a decreased capability to antagonize innate immunity, ultimately leading to the attenuated phenotype of the octamer mutant MHVoctm in mice. Consequently, because (i) the levels of viral titers and RNA were not decreased with the time (Fig. 5F and H) and (ii) MHVoctm could express viral protein in the livers of mice, the results suggest that MHVoctm can replicate in mice (Fig. 5G). In addition, because the levels of viral titers, proteins and viral RNA were also lower in MHVoctm-infected mice than in MHVwt-infected mice, the results suggest that MHVoctm is attenuated in mice. Since MHVoctm with the mutated octamer can replicate well in cell cultures (Fig. 1) but is attenuated in mice (Fig. 5), the results together suggest that the octamer-mutated MHVoctm is a potential vaccine candidate.
Fig. 5
MHVoctm with octamer mutation is attenuated in mice. (A) Timeline for physical examination of the phenotype of MHVwt- or MHVoctm-infected mice, including body weight (B), clinical score (C) and survival rate (D). 106 pfu MHVwt or MHVoctm were used for the experiment. (E) Timeline for physical examination of virus titer (F), translation efficiency (G), synthesis of viral RNA (H) and histopathology (I) in MHVwt- or MHVoctm-infected mice. Histopathology examination (scale bar = 100 μm) revealed coagulation necrosis and vacuolated hepatocytes in the livers of mice inoculated with MHVwt but not with MHVoctm. The uncropped gels for (G) were shown in Figure S2. dpi, days postinfection; MHVwt or wt, wild-type MHV; MHVoctm or octm, MHV with octamer mutation. Statistical significance was evaluated using a t test: ** P < 0.01, *** P < 0.001, **** P < 0.0001
MHVoctm can be used as an attenuated vaccine to protect mice against MHVwt infectionSince the octamer mutant MHVoctm can grow to high titers in cultured cells and is attenuated in mice (Figs. 1 and 5), the octamer mutant MHVoctm has the great potential as a vaccine candidate against MHVwt infection. For this purpose, 3-week-old male BALB/c mice were inoculated with DMEM or 106 pfu of MHVoctm once or twice followed by challenge with 106 pfu of MHVwt (Fig. 6A). Before the challenge with MHVwt, significant weight loss, clinical signs and death were not observed in all groups. After the challenge with MHVwt, weight loss, clinical signs and death were also not observed in mice inoculated with octamer mutant MHVoctm once or twice (Fig. 6B-D). In contrast, mice inoculated with DMEM started to show significant weight loss and clinical signs after 1 day of challenge and started to die after 2 days of challenge (Fig. 6B-D). In addition, the viral RNA detected from mice inoculated with octamer mutant MHVoctm once or twice was significantly less than that inoculated with DMEM (8.2 × 101/ng vs. 4.3 × 108/ng, Fig. 6E). By histopathological examination, coagulation necrosis and vacuolated hepatocytes (ballooning degeneration) were observed in the livers of mice inoculated with DMEM followed by challenge with MHVwt (DDW group) but not in groups of mice inoculated with octamer mutant MHVoctm once (DOW group) or twice (OOW group) followed by challenge with MHVwt (Fig. 6F). Based on the results of weight loss, clinical signs, survival rate, viral RNA detection and histopathological examination, manipulation of the conserved octamer is a promising strategy for vaccine design against coronavirus infection.
Fig. 6
Examination of the safety and efficacy of the attenuated vaccine candidate MHVoctm. (A) Timeline and experimental groups for examination of the safety and efficacy of the attenuated vaccine MHVoctm. A total of 106 pfu of MHVwt or MHVoctm were used for the experiment. (B)-(F) Examination of body weight (B), clinical score (C), survival rate (D), synthesis of viral RNA (E) and pathological changes in which coagulation necrosis and vacuolated hepatocytes (ballooning degeneration) were observed in the livers of mice inoculated with DMEM wt but not with MHVoctm after challenge with MHVwt (scale bar = 100 μm) (F). dpc, days postchallenge; MHVwt, wild-type MHV; MHVoctm, MHV with octamer mutation. Statistical significance was evaluated using a t test: *** P < 0.001, **** P < 0.0001
Refinement of the vaccination strategy for the attenuated vaccine candidate MHVoctm by reducing the dosage and times of vaccinationTo further determine the vaccine potential of the mutant MHVoctm, the vaccination strategy was refined by reducing the times and the virus dosage for vaccination. For this purpose, mice were inoculated with the octamer mutant MHVoctm only once, with 102, 104 or 106 pfu of virus (Fig. 7A). As shown in Fig. 7B-D and F, no significant weight loss, clinical signs, death or histopathological changes were observed in mice inoculated with 102, 104 or 106 pfu of octamer mutant MHVoctm followed by challenge with MHVwt. In addition, a much lower amount of viral RNA was detected in mice inoculated with 102, 104 or 106 pfu of octamer mutant MHVoctm followed by challenge with MHVwt in comparison with that in mice inoculated with DMEM followed by challenge with MHVwt. (8.1 × 101/ng vs. 1.5 × 106/ng, Fig. 7E).
Fig. 7
The safety and efficacy of the attenuated vaccine candidate MHVoctm with modified vaccination strategy. (A) Timeline and experimental groups for examination of the efficacy and safety of the attenuated vaccine MHVoctm. The reduced dosages (106, 104 and 102 pfu) of MHVoctm and one vaccination were used for the experiment. (B)-(F) Examination of body weight (B), clinical score (C), survival rate (D), synthesis of viral RNA (E) and pathological changes in which coagulation necrosis and vacuolated hepatocytes (ballooning degeneration) were observed in the livers of mice inoculated with DMEM but not with MHVoctm after challenge with MHVwt (scale bar = 100 μm) (F). dpc, days postchallenge; MHVwt, wild-type MHV; MHVoctm, MHV with octamer mutation. Statistical significance was evaluated using a t test: * P < 0.05
To examine whether MHVoctm was able to elicit neutralizing antibody production, a serum virus neutralization assay was performed. As shown in Fig. 8 and S3, neutralizing antibodies were measured, and the average titer increased moderately in a dose-dependent manner. In addition, booster vaccination induced more neutralizing antibody production than only one vaccination. The results suggest that the octamer mutant MHVoctm can induce neutralizing antibody production.
Taken together, the results suggest that inoculating mice with 102 pfu of the octamer mutant MHVoctm one time is sufficient to provide protection against MHVwt infection, thus further demonstrating that manipulation of conserved octamers is a novel and promising strategy to develop a live attenuated vaccine against coronavirus infection.
Fig. 8
Titers of serum virus neutralization antibodies in vaccinated mice. (A) Timeline and experimental groups for the measurement of neutralizing antibodies. (B)-(D) Examination of body weight (B), clinical score (C) and survival rate (D). (E) Titers of neutralizing antibodies. dp2i, days post-2nd infection
留言 (0)