
記住我
As one of the most common neurodegenerative disorders impairing the elderly population, Alzheimer’s disease (AD) is typically characterized by cognitive dysfunction and will cause great burden to patients and caregivers. Based on the results of numerous studies, intracellular neurofibrillary tangles (NFTs) and extracellular senile plaques (SPs) within the hippocampus have been conceived as the two major pathogenic mechanisms initiating AD [1]. Generally, both proteins are formed by the dysfunction of specific neural proteins, such as the misfolding of highly phosphorylated Tau protein and the deposition of amyloid-beta (Aβ) peptides [1], which eventually leads to a cascade of neurodegeneration during the progression of AD [2]. Due to the increase in lifetime during the last decade worldwide, the number of AD patients has been steadily rising, and despite significant efforts to create disease-modifying treatments for AD treatment, conventional pharmacotherapy has so far been unable to stop or reverse the progression of AD disease. In such situation, investigating new therapeutic targets or managing strategy for treating AD is not only warranted but also essential.
Traditional Chinese Medicine (TCM) is a treasury for discovering natural compounds with multiple promising pharmacological effects. Certain natural compounds from TCM have been shown to ameliorate cognitive impairment by improving mitochondrial dysfunction and repairing neuroplasticity in patients with neurodegenerative disorders via multiple mechanisms [3–5]. Regarding the treatment of AD, there are some Chinese medicine extracts also showing considerable effects. For instance, hederagenin, a triterpene compound, isolated from Matoa (Pometia pinnata) fruit can improve AD model through PPARα/TFEB-mediated autophagy [6]. Ginsenoside Rb1 also shows neuroprotective effects in a mouse model of AD by suppressing the deposition of Aβ and the formation of phosphorylated tau protein [7]. Cynomorium songaricum Rupr. is a TCM herb and has been widely employed for the treatment of sexual disorders, such as erectile dysfunction and menstrual problems [8]. With increasing attention paid to the neurogenerative effects of TCM, the neuroprotective potential of C. songaricum was also tested. As reported by Cheng et al., extract of C. songaricum can ameliorate mitochondrial dysfunction in in-vitro models of AD [9]. Additionally, the extract from C. songaricum can also alleviate memory impairments by activating CREB/BDNF axis in ovariectomized rats [10]. Based on the previous study by Meng et al., the functional compounds mediating the bioactivity of C. songaricum are majorly flavonoids such as epicatenchin, isoquercitrin, and phlorizin [9]. Flavonoids are well-characterized health-benefiting agents due to their high antioxidant capacity as well as their anti-allergic, anti-inflammatory, anti-microbial, and anti-cancer activities [11]. Moreover, this type of compound can also contribute to the attenuation of neurodegenerative disorders such as AD and PD [12–15]. Therefore, the neuroprotective effects of C. songaricum extract may be also attributed to the functions of flavonoids.
To verify the hypothesis, the current study employed total flavonoids of C. songaricum to treat a rat AD model induced by the injection of Aβ1-42. The effects of C. songaricum flavonoid extract (CS) on the cognitive function, brain histology, and generation of neurotransmitters were detected to explicitly demonstrate the neuroprotective effects of CS. Additionally, to preliminarily explain the pathways mediating the function of CS, the current study also detected the changes in BDNF/TrkB axis. It is well-known that the function of BDNF via stimulating its high-affinity receptor TrkB contributes to cell differentiation, cell survival, synaptic plasticity, and both embryonic and adult neurogenesis in central nervous system (CNS) [16]. Previous studies have also demonstrated possible involvement of altered expression and action of the BDNF in the pathogenesis of AD due to an impaired neurogenesis during aging [16]. Based on the results of these assays, we attempted to reveal the role and associated mechanism of total flavonoids in the neuroprotective effects of C. songaricum.
Methods Chemicals and agentsAβ1-42 (No. 04010011827, Qiangyao Bio) was incubated at 37 °C for one week prior to injection and was then lyophilized. 1 mg Aβ1-42 lyophilized powder was dissolved in 50 μl of DMSO and diluted to 5 μg/μl with normal saline and kept in a -80 °C refrigerator for subsequent assays. The CS were prepared routinely following our previously published work [17] and mixed with normal saline into solutions into concentrations of 2.5, 5, and 10 mg/ml. Donepezil hydrochloride (5 mg/tablet) was purchased from Eisai Pharmaceutical (No. 2010023, China). TUNEL kit was purchased from Roche (No. 49330900, Switzerland); BCA protein concentration determination kit was purchased from Beyotime (No. P0009, Shanghai, China). Antibody against Bax was obtained from Proteintech Group (No. 50599-2-1g, Wuhan, China); antibodies against Bcl-2 (No. A19693), caspase-9 (No. A18676), and TrkB (No. A19832) were obtained from ABclonal (Wuhan, China); antibody against BDNF (No. Ab108319) Abcam (Shanghai, China); antibody against caspase-3 (No. AF6311) was purchased from Affinity Biosciences LTD (Jiangsu China). ELISA kits for detecting choline acetyltransferase (ChAT) (No. ZC-36507), acetylcholine (ACH) (No. ZC-37533), and acetylcholinesterase (AChE) (No. ZC-37535) were purchased from Zcibio (Shanghai, China).
Animals and AD model inductionMale Wistar rats (4 months old and weighed 250 ± 50 g) were provided by Lanzhou Veterinary Research Institute of the Chinese Academy of Agricultural Sciences Animal and all the animal assays were performed under the approval of Laboratory of the Gansu Institute of Drug Inspection and Investigation (approval no. SYXK (Gan) 2020-0006). After one week of adaptive feeding with normal diets in our lab, 70 rats were randomly assigned to Control group, Sham group, AD group, Donepezil group, CSL (CS low) group, CSM (CS Medium) group, and CSH (CS High) group (10 for each group). For AD model, Donepezil, CSL, CSM, and CSH groups, rats underwent intraperitoneal injection of 35 mg/kg 3% pentobarbital sodium to induce anesthesia and the location of rats’ hippocampus was established using brain stereotaxic coordinates [18]: 3.0 mm posterior to the bregma, 2.2 mm lateral to each side, and 3 mm deep below the dura mater. Then a hole at the designated site was drilled using a dental drill, and 1 μl of Aβ1-42 solution (5 μg/μl) was injected slowly (0.2 μl/min) on each side using a microinjector. For rats in the Control group, no treatment was performed, while for rats in Shzam surgery group, the injection would be replaced by an equal volume of normal saline. After withdrawing the needle, the wound was sutured and the rats were singly housed until the wound was fully healed. 3 days after the modeling, rats in Donepezil group were gavaged with a Donepezil solution at a dose of 0.5 mg/kg once a day for 28 consecutive days, and rats in CSL, CSM, and CSH groups were given CS solutions at doses of 25, 50, and 100 mg/kg once a day for 28 consecutive days, respectively. For rats in Control group, Sham group, and AD group, an equivalent volume of normal saline was given by gavage once a day for 28 consecutive days.
Morris water maze testThe Morris water maze (MWM) was utilized to assess the learning and memory abilities of the rats. The assays were conducted according to previously reported procedures [19,20] by two investigators who were blind to the animal group. The test consisted of a 4-day visible platform trial and a 1-day probe trial. During the visible platform trial, the rats were allowed to swim for 60 s to find the platform four times a day. If a rat failed to locate the platform, the investigators would assist the rat to arrive and remain on the platform for 10 s before proceeding to the next test. On the 5th day, the probe trial was conducted with the platform removed. The rats were given 60 s to explore the area where the platform was previously located, and the time spent and number of entries through each quadrant of the former platform position were measured by the investigators.
Step-down testThe step-down test was performed to evaluate the cognitive and memory functions of the rats [21,22], and was divided into a learning phase and a memory phase. The step-down test was performed with a Step-down Recorder (DB097, ShuduoBao, Beijing, China). During the learning phase, the rats were allowed to freely explore the platform box for 3 min, and then a 36 V electric shock was applied. The correct response for the rat after receiving the stimulation was to jump onto the platform to avoid the shock, while the incorrect response was to jump off the platform after the rats had already been on the platform. The memory retention ability of the rats was tested 24 h later. During memory phase, the electronic shock was maintained for five minutes, and the latency to jump onto the platform for the first time and the number of incorrect responses within 5 min were recorded. Upon completion of the step-down test, rats were anesthetized with 3% pentobarbital sodium for the collection of the femoral artery blood. Then the rats were sacrificed with overdose pentobarbital sodium and hippocampus tissues were collected and then subjected to investigation of H&E staining, TUNEL 427 staining, and TEM.
Histological detectionThe hippocampus tissues were subjected to H&E staining to determine the histological changes induced by Aβ1-42 injection. The tissues were sectioned, dewaxed, rehydrated, and stained with hematoxylin and eosin. The images were captured an Olympus microscope (Olympus, Japan) at ×400 magnification.
Neuron apoptosis was detected with TUNEL staining: briefly, after dehydration, paraffin embedding, and deparaffinization, the fixed brain tissue was subjected to an 8-minute microwave recovery with citrate buffer. Then TUNEL incubation solution was added and were incubated with the samples at 37 °C for 1 h. Afterwards, DAPI staining was incubated with the samples for 15 min to stain the nuclei, followed by mounted with glycerin gelatin. The apoptosis rate was calculated following apoptotic cells/total cells × 100% using a digital slice scanner.
Ultrastructure of hippocampus tissues was determined via TEM detection of autophagosome: cells were embedded in Epon resin and morphology of autophagosome was detected using a transmission electron microscope (JEM-1400FLASH, Japan) at an acceleration voltage of 120 kV at 40 000× magnification.
Immunochemical detectionThe expression level and distribution of Aβ1-42 in hippocampus tissues were determined with immunochemical (IHC) detection. Tissues were fixed with glutaraldehyde, sectioned, dewaxed with dimethylbenzene, and hydrated with alcohol. Then the primary antibody against Aβ1-42 (1:1000) was incubated with the tissues at 4 °C overnight, followed by the incubation by the secondary antibody (1:2000) (ab288151, Abcam, China) at 37 °C for 30 min. After re-stained with hematoxylin and dehydrated, the images were scanned and captured using an Aperio ScanScope GL (Aperio Technologies, Vista, CA, USA) at 400× magnification.
ELISA detectionThe production of ChAT, ACH, and AChE was detected following the manufacturers’ instruction.
Western blottingTotal protein was extracted from hippocampal tissues using Protein lysis buffer (G2002-100ML, Service-Bio, Wuhan, China) and protein concentration was determined using the BCA protein quantification kit. 30 μg of protein was loaded and electrophoresed at 100 V for 15 min. The membrane was then transferred at 200 mA for 1 h, blocked with 5% skimmed milk, and incubated with primary antibodies against Bax (1:5000), Bcl-2 (1:2000), BDNF (1:2000), caspase-3 (1:2000), caspase-9 (1:2000), TrkB (1:2000), and β-actin (1:100 000) at 4 °C overnight. After washing, the membrane was incubated with secondary antibodies, and the protein bands were visualized using an ultra-sensitive ECL chemiluminescence kit. The gel imaging system was used to scan the bands, and the relative expression of the target protein was calculated with β-actin as an internal reference.
Statistical analysisAll the data was represented as mean ± SD and the difference among groups was analyzed using one-way ANOVA followed by Tukey’s test. All the statistical analyses were performed with IBM SPSS 21.0 with a significant level of 0.05 (two-tailed P value).
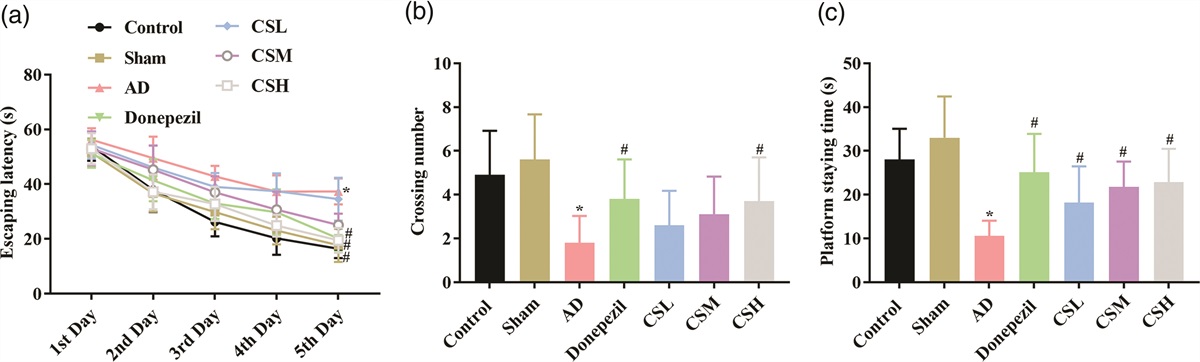
Results CS improved the cognitive functions of AD rats in both MWM and step-down testsThe effects CS on the cognitive functions of AD were assessed with MWM and step-down tests. The positioning navigation experiment in the MWM test reflected the learning and memory abilities of rats: the fluctuation of the data suggests that the average escaping latency decreases with the increase of testing days, especially from third to fifth days, indicating that Aβ1-42 reduced the learning and memory ability of rats. During the first to fifth days there was no significant difference in the average escaping latency of rats in Control group and Sham group (P > 0.05) (Fig. 1a). Compared with Sham group, the average escaping latency of rats in AD group was significantly prolonged (P < 0.05) (Fig. 1a). The average escaping latency of rats in Donepezil group was significantly shortened (P < 0.05). Regarding the effects of CS, the extract exerted the cognitive improving effects in a dose-dependent manner. CSL had no significant effects on the cognitive function of AD rats, while the average escape latency of rats in the CSM group was significantly shortened during third to fifth days compared with AD group (P < 0.05), and the functioning time course for CSH was from the second to fifth days (P < 0.01) (Fig. 1a).
 Fig. 1:
Fig. 1: CS improved the memory and learning ability of AD rats in MWM test. Rats were subjected to transcranial injection of Aβ1-42 and handled with CS of three doses. The memory and learning ability of rats in different groups was then assessed with MWM test. (a) Analysis of escaping latency during the 4-day visible platform trial. (b) Analysis of crossing number through the platform location during the 1-day probe trial. (c) Analysis of staying time at the platform location during the 1-day probe trial. ‘*’ P < 0.05 vs. Sham group. ‘#’ P < 0.05 vs. AD group.
On the sixth day of the experiment, the platform was removed to test spatial memory ability of the rats. Compared with Control group, there was no significant difference in the number of platform crossings and target quadrant dwell time of rats in Sham group (P > 0.05) (Fig. 1b and 1c). However, the number of platform crossings and target quadrant dwell time of rats in the model group were significantly reduced in AD group compared with Sham group (P < 0.05) (Fig. 1b and 1c). The number of platform crossings and target quadrant dwell time were then restored in rats administrated with donepezil and high-dose CS (P < 0.05) (Fig. 1b and 1c), and the target quadrant dwell time of rats in CSL and CSM groups were significantly increased (P < 0.05) (Fig. 1c), indicating that Aβ1-42 weakened the spatial exploration ability of rats, while CS restored the ability in a dose-dependent manner.
Similar results were also detected with step-down tests: compared with Sham group, rats in AD group had a significantly shorter latency in the learning and memory phase (P < 0.05) (Fig. 2a) and a significant increase in the number of errors (P < 0.05) (Fig. 2b). Regarding rats in Donepezil group and CSH group, a significantly prolonged latency in the learning and memory phase (P < 0.05) and a significant decrease in the number of errors (P < 0.05) were recorded (Fig. 2), further confirming the improving effects of CS on the cognitive function of AD rats.
 Fig. 2:
Fig. 2: CS improved the memory and learning ability of AD rats in step-down test. Rats were subjected to transcranial injection of Aβ1-42 and handled with CS of three doses. The memory and learning ability of rats in different groups was then assessed with step-down test. (a) Analysis of latency before jump. (b) Analysis of error number before jump. ‘*’ P < 0.05 vs. Sham group. ‘#’ P < 0.05 vs. AD group.
CS improved the brain tissue destruction and neuron apoptosis in the hippocampus tissue of AD ratsThe effect of CS on the hippocampal histology of AD rats was firstly detected with H&E staining. As shown in Fig. 3, pyramidal cell apoptosis along with blurred cell structure, increased cytoplasmic staining, unclear nuclear boundaries, and disordered arrangement of some pyramidal cells were observed in AD group. The impairments on hippocampus tissues were dramatically improved by donepezil and CS: fewer apoptotic pyramidal cells and ordered arrangement of pyramidal cells could be observed for the four groups, indicating the repairment of hippocampus tissues by CS (Fig. 3). Additionally, the effects of CS on histology of hippocampus tissues were also exerted in a dose-dependent manner: in CSH group, the brain tissues had intact soft meningeal structure, and no obvious inflammatory exudation and no neuronal degeneration, could be observed (Fig. 3). Similar results were also detected for TUNEL staining and TEM observation: the number of TUNEL-positive neurons were firstly increased by Aβ1-42 injection and then suppressed by CS in a dose-dependent manner (Fig. 3 and Figure S1, Supplemental digital content 1, https://links.lww.com/WNR/A716), and the ultrastructure of hippocampus tissues was also improved by CS by restoring synapse number, repairing neuron morphology, and suppressing neuron apoptosis (Fig. 3).
 Fig. 3:
Fig. 3: CS ameliorated Aβ1-42-induced brain structure destruction and tissue apoptosis in AD rats. Hippocampus tissues were collected and then subjected to investigation of H&E staining, TUNEL staining, and TEM. Green arrow, apoptotic pyramidal cells; Black arrow, dark neurons; Blue arrow, disordered pyramidal cells; Red arrow, destructed cell structure; Yellow arrow, deep cytoplasm; Green circle, regular synapses; Mi, mitochondrial; Yellow circle, increased distance between synapses; Red circle, dissolved synaptic vesicles; Black circle.
CS increased the production of ChAT, ACH, while decreased the production of AChE in the serum and hippocampus, and inhibited the generation of Aβ1-42 in the hippocampus of AD ratsThe production ChAT, ACH, and AChE was detected with ELSIA assays both in serum and hippocampus tissues of rats. In hippocampus tissues, the levels of ChAT and ACH were significant suppressed by Aβ1-42 injection (Fig. 4a and 4b), while the level of AChE was induced (Fig. 4c). Similar results were also detected in serum samples (Fig. 4d–4f). The producing pattern of the markers was then restored by donepezil and CS, and the effects of CS were exerted in a dose-dependent manner (Fig. 4a–4f). Regarding the production and distribution of Aβ1-42, the establishment of AD model increased the production and distribution of the protein in the hippocampus of rats, which was then restricted by CS in a dose-dependent manner (Fig. 4g).
 Fig. 4:
Fig. 4: CS increased the production of ChAT, ACH, decreased the production of AChE and the generation of Aβ1-42 in AD rats. The production of ChAT, ACH, and AChE in serum and hippocampus samples was detected with ELISA. The production and distribution of Aβ1-42 was detected with IHC. (a) Analysis of ChAT level in hippocampus tissues. (b) Analysis of ACH level in hippocampus tissues. (c) Analysis of AChE level in hippocampus tissues. (d) Analysis of ChAT level in serum samples. (e) Analysis of ACH level in serum samples. (f) Analysis of AChE level in serum samples. G, images and analysis of Aβ1-42 level in hippocampus tissues. ‘*’ P < 0.05 vs. Sham group. ‘#’ P < 0.05 vs. AD group.
CS activated BDNF/TrkB axis while inhibited apoptosis-related pathways in the hippocampus of AD ratsThe potential mechanism underlying the function of CS was explored by detecting changes in BDNF/TrkB axis. The expression levels of BDNF and TrkB were both suppressed by Aβ1-42 injection, which was associated with upregulation of pro-apoptosis markers such as Bax, caspase-3, and caspase-9 and downregulation of anti-apoptosis marker Bcl-2 (Fig. 5). The expression patterns of these markers were then reversed by donepezil and CS, indicating that the neuro-protective effects of CS might be related to the activation of BDNF/TrkB axis. Additionally, the effects of CS on the activity of signaling transduction were also exerted in a dose-dependent manner with high dose of CS showing the strongest effects (Fig. 5).
 Fig. 5:
Fig. 5: CS increased the expression of BDNF, TrkB, and Bcl-2, while inhibited caspase-9, caspase-3, and Bax in hippocampus tissues of AD rats. The expression levels of BDNF, TrkB, Bcl-2, caspase-9, caspase-3, and Bax in hippocampus tissues were detected with western blotting. ‘*’ P < 0.05 vs. Sham group. ‘#’ P < 0.05 vs. AD group.
DiscussionC. songaricum is a Chinese herbal medicine commonly employed for the treatment of sexual disorders [8] by modifying the kidney, enriching essence and blood, and promoting bowel movement, and also has actions consistent with the TCM theory of the initiation of AD. For instance, the study by Cheng et al. showed that extracts from C. songaricum can protect neurons against AD symptoms in vitro by ameliorating mitochondrial ultrastructure impairments and dysfunction [9]. In the study by Tian et al., the authors declare that extracts from C. songaricum can attenuate memory impairment by activating CREB/BDNF pathway via suppression of p38MAPK/ERK pathway [10]. These previous studies demonstrate that the neuroprotective effects of C. songaricum can be exerted via multiple mechanisms working together, including reducing oxidative stress levels, removing oxygen free radicals, regulating mitochondrial dynamics imbalance, promoting cell regeneration and synaptic growth, protecting the morphology of hippocampal neurons in brain tissues [23], which finally leads to the improved learning and memory abilities. However, most previous studies employed extracts of C. songaricum as the treating agent, few studies have provided valuable information for determining the major functional components mediating the neuroprotective function of C. songaricum, which limited the further application and development of products of C. songaricum.
According to the investigation by Meng et al., the major functional compound family mediating the bioactivity of C. songaricum are flavonoids such as epicatenchin, isoquercitrin, and phlorizin [9]. In the current study, we attempted to evaluate the treatment potential of CS against AD model. Based on our data, CS could improve cognitive function, repair brain tissue structure, and suppress neuron apoptosis in rats injected with Aβ1-42, and the effects were associated with the increased production of ACH, ChAT, decreased the production of AChE as well as the inhibited generation of Aβ1-42. ACH is a key neurotransmitter and its level is positively correlated with learning and memory abilities [24]. The balance of ACH is generally modulated by AchE and ChAT. However, during the progression of AD, a significant decrease in ACH synthesis, storage, and release was initiated, which in turn causes learning, memory, and recognition dysfunction [25]. Therefore, the changes regarding these phenotypes and levels of neurotransmitters in the current study solidly demonstrated the anti-AD function of CS.
As a class of well-characterized bioactivity components, flavonoids have shown protective effects against diverse diseases such as atherosclerosis and cardiovascular diseases via its anti-oxidant and anti-inflammatory activities [26]. Regarding the effects against neurogenerative disorders, multiple studies have also reported the treatment effects of flavonoids against disorders such as AD and PD. For instance, the study by Andrikopoulos et al. demonstrates that epigallocatechin gallate function as an anti-amyloidosis and antioxidation agent [12], and the study by Gao et al. shows that cyanidin-3-O-glucoside can alleviate AD syndromes by disturbing the aggregation of single Aβ peptide [13]. The protective effects of other flavonoids such as quercetin and oleuropein against neurogenerative disorders are also reported [14,15]. The current study formed a supplement to these previous reports and connected the neuroprotective effects of C. songaricum with flavonoids, which not only provides the valuable information for explaining the bioactivity of C. songaricum, but also guides the future development of anti-AD strategy with C. songaricum.
In addition to assessing the effects of CS on AD symptoms, the current study also attempted to reveal the molecular pathways mediating the function of CS. Thus, the changes in Aβ1-42-induced changes in BDNF/TrkB and apoptosis-related pathways were also detected. The administration of CS restored the activity of BDNF/TrkB axis and suppressed the activation of pro-apoptosis pathways. BDNF is one of the most widely distributed neurotrophic factors in the mammalian hippocampus and has multiple biological activities, such as promoting nervous system development, protecting against neuronal damage, and improving learning and memory abilities [27]. Regarding its role in the development of AD, the level of BDNF is substantially decreased in the hippocampus, olfactory cortex, neocortex, and basal forebrain of AD patients [28]. In the current study, the suppression of BDNF was induced by Aβ1-42, the neurotoxicity of which is considered to be the major causable factor contributing to the synaptic and cognitive dysfunction associated with AD [29]. The inhibited expression of BNDF subsequently blocked the phosphorylation of TrkB as well as the activation of downstream pathways such as PI3K/Akt, finally leading to the impaired memory and learning ability of AD patients [30]. The impaired activation of PI3K/Akt will also increase the expression of pro-apoptosis molecules such as Bax, caspase-3, and caspase-9, while suppressed the expression of anti-apoptosis molecules such Bcl-2, contributing to the increased apoptosis in brain neurons and exacerbating the pathogenesis of AD [31]. Thus, the restored activity of BNDF/TrkB axis by CS might explain the mechanism underlying the anti-AD function of CS: the activated BNDF/TrkB axis by CS modulates the activity of downstream effectors such as PI3K/Akt, Bcl-2, Bax, and caspases, and contributes to the attenuation of tissue destruction and restored production of neurotransmitters, which lead to the improved cognitive function of AD rats.
Collectively, CS can improve learning and memory abilities in Aβ1-42-induced AD model rats. The effects may depend on the activation the hippocampal BDNF/TrkB signaling pathway, which will increase ACH level, inhibit neuron apoptosis, and increase synaptic plasticity. However, the current study only provides a preliminary explanation to the anti-AD function of CS. The exact composition of CS is not elaborated by the current study, and the changes in the downstream of BNDF/TrkB axis also needs further exploration. Thus, more comprehensive studies will be performed in our lab to promote the application of C. songaricum in the management of AD and other neurodegenerative disorders.
AcknowledgementsThis work was supported by Gansu Provincial Youth Science and Technology Fund (No.21JR7RA634) and Gansu Provincial Natural Science Fund (No. 20JR5RA154).
ZRG performed data curation, formal analysis, and wrote the draft of MS. XL and YG performed data curation. MQ and BG performed conceptualization and revised the MS.
Data will be provided on reasonable request.
Conflicts of interestThere are no conflicts of interest.
References 1. Eckert A, Schulz KL, Rhein V, Götz J. Convergence of amyloid-beta and tau pathologies on mitochondria in vivo. Mol Neurobiol 2010; 41:107–114. 2. Coleman PD, Yao PJ. Synaptic slaughter in Alzheimer’s disease. Neurobiol Aging 2003; 24:1023–1027. 3. Zhao J, Yang J, Ding L, Wang F, Lin L. A Review of the Pathogenesis and Chinese Medicine Intervention of Alzheimer’s Disease. J Integr Neurosci 2022; 22:2. 4. Liao Y, Wang X, Huang L, Qian H, Liu W. Mechanism of pyroptosis in neurodegenerative diseases and its therapeutic potential by traditional Chinese medicine. Front Pharmacol 2023; 14:1122104. 5. Du H, Yan SS. Mitochondrial medicine for neurodegenerative diseases. Int J Biochem Cell Biol 2010; 42:560–572. 6. Xie ZS, Zhao JP, Wu LM, Chu S, Cui ZH, Sun YR, et al. Hederagenin improves Alzheimer’s disease through PPARα/TFEB-mediated autophagy. Phytomedicine 2023; 112:154711. 7. Shalaby AM, Alnasser SM, Ahmed Khairy D, Alabiad MA, Alorini M, Jaber FA, et al. The neuroprotective effect of ginsenoside Rb1 on the cerebral cortex changes induced by aluminium chloride in a mouse model of Alzheimer’s disease: A histological, immunohistochemical, and biochemical study. J Chem Neuroanat 2023; 129:102248. 8. Cheng D, Zheng J, Ma S, Murtaza G, Wahab A, Yu C, et al. Chemical constituents, and pharmacological and toxicological effects of Cynomorium songaricum: an overview. Trop J Pharm Res 2017; 16:2689–2696. 9. Cheng D, Su L, Wang X, Li X, Li L, Hu M, et al. Extract of Cynomorium songaricum ameliorates mitochondrial ultrastructure impairments and dysfunction in two different in vitro models of Alzheimer’s disease. BMC Complement Med Ther 2021; 21:206. 10. Tian FZ, Chang HS, Liu JX, Zheng J, Cheng D, Lu Y. Cynomorium songaricum extract alleviates memory impairment through increasing CREB/BDNF via suppression of p38MAPK/ERK pathway in ovariectomized rats. Evid Based Complement Alternat Med 2019; 2019:9689325. 11. Ohtsuki T, Akiyama J, Shimoyama T, Yazaki S, Ui S, Hirose Y, et al. Increased production of antioxidative sesaminol glucosides from sesame oil cake through fermentation by Bacillus circulans strain YUS-2. Biosci Biotechnol Biochem 2003; 67:2304–2306. 12. Andrikopoulos N, Li Y, Nandakumar A, Quinn JF, Davis TP, Ding F, et al. Zinc-epigallocatechin-3-gallate network-coated nanocomposites against the pathogenesis of amyloid-beta. ACS Appl Mater Interfaces 2023; 15:7777–7792. 13. Gao J, Fu J, Gao X, Yang D. Molecular mechanism of cyanidin-3-o-glucoside disassembling aβ fibril in silico. Nutrients 2023; 15:1–8. 14. Rafiq H, Farhan M, Rafi H, Rehman S, Arshad M, Shakeel S. Inhibition of drug induced Parkinsonism by chronic supplementation of quercetin in haloperidol-treated wistars. Pak J Pharm Sci 2022; 35:1655–1662. 15. Singh R, Zahra W, Singh SS, Birla H, Rathore AS, Keshri PK, et al. Oleuropein confers neuroprotection against rotenone-induced model of Parkinson’s disease via BDNF/CREB/Akt pathway. Sci Rep 2023; 13:2452. 16. Numakawa T, Odaka H. Brain-derived neurotrophic factor and neurogenesis. Factors Affecting Neurodevelopment: Elsevier; 2021. pp. 121–131. 17. Lyv X, Gu ZR, Qi M, Zhang R, Guo Y, Mao XW, et al. Optimization of extraction and purification process and adsorption characteristics of cynomorium songaricum flavonoids. Chin Wild Plant Res 2022; 41:41–49. 18. Paxinos G, Watson C. The rat brain in stereotaxic coordinates: hard cover edition. Elsevier; 2006. 19. Chu Q, Jian-Chun YU. Improvement of acupuncture on cognitive function in senescence accelerated mouse P8. Chin J Behav Med Sci 2005; 14:964–965. 20. Liu CZ, Yu JC, Cheng HY, Jiang ZG, Li T, Zhang XZ, et al. Spatial memory performance and hippocampal neuron number in osteoporotic SAMP6 mice. Exp Neurol 2006; 201:452–460. 21. Zhu X, Cai L, Liu J, Zhu W, Cui C, Ouyang D, et al. Effect of seabuckthorn seed protein and its arginine-enriched peptides on combating memory impairment in mice. Int J Biol Macromol 2023; 232:123409. 22. Zhang Y, Huang H, Yao C, Sun X, He Q, Choudharyc MI, et al. Fresh Gastrodia elata Blume alleviates simulated weightlessness-induced cognitive impairment by regulating inflammatory and apoptosis-related pathways. Front Pharmacol 2023; 14:1173920. 23. Meng HC, Wang S, Li Y, Kuang YY, Ma CM. Chemical constituents and pharmacologic actions of Cynomorium plants. Chin J Nat Med 2013; 11:321–329. 24. Wang Q, Wang C, Shu Z, Chan K, Huang S, Li Y, et al. Valeriana amurensis improves Amyloid-beta 1-42 induced cognitive deficit by enhancing cerebral cholinergic function and protecting the brain neurons from apoptosis in mice. J Ethnopharmacol 2014; 153:318–325. 25. Ferreira-Vieira TH, Guimaraes IM, Silva FR, Ribeiro FM. Alzheimer’s disease: targeting the cholinergic system. Curr Neuropharmacol 2016; 14:101–115. 26. Lotito SB, Frei B. Consumption of flavonoid-rich foods and increased plasma antioxidant capacity in humans: cause, consequence, or epiphenomenon? Free Radic Biol Med 2006; 41:1727–1746. 27. Kowiański P, Lietzau G, Czuba E, Waśkow M, Steliga A, Moryś J. BDNF: a key factor with multipotent impact on brain signaling and synaptic plasticity. Cell Mol Neurobiol 2018; 38:579–593. 28. Kim BY, Lee SH, Graham PL, Angelucci F, Lucia A, Pareja-Galeano H, et al. Peripheral brain-derived neurotrophic factor levels in alzheimer’s disease and mild cognitive impairment: a comprehensive systematic review and meta-analysis. Mol Neurobiol 2017; 54:7297–7311. 29. Amidfar M, de Oliveira J, Kucharska E, Budni J, Kim YK. The role of CREB and BDNF in neurobiology and treatment of Alzheimer’s disease. Life Sci 2020; 257:118020. 30. Barage SH, Sonawane KD. Amyloid cascade hypothesis: pathogenesis and therapeutic strategies in Alzheimer’s disease. Neuropeptides 2015; 52:1–18. 31. Cui J, Shan R, Cao Y, Zhou Y, Liu C, Fan Y. Protective effects of ginsenoside Rg2 against memory impairment and neuronal death induced by Aβ25-35 in rats. J Ethnopharmacol 2021; 266:113466.
留言 (0)