
記住我








The term laser is an acronym for light amplification by the stimulated emission of radiation. Since the first laser was developed in 1960, the role that lasers play in various fields, including biology, chemistry, and medicine, has increased steadily [1]. Many procedures have only become possible with the use of lasers. Currently, more than forty different types of lasers have been found in medicine [2]. In the early 1970s, lasers were introduced into the field of gynecology for surgery, excision and ablation of tissue [3,4,5]. In the 1980 and 1990 s, lasers began to be used in infertility treatment of endometriosis, tubal surgery, ectopic pregnancy and polycystic ovarian syndrome through operative microscopes and laparoscopes [6,7,8]. Since the successful establishment of assisted reproductive technology (ART), in vitro manipulation of gametes, zygotes and embryos has been an essential integral part of ARTs. Laser technology was introduced into the field of ART as a valuable tool to replace many mechanical and chemical procedures used in the in vitro manipulation of gametes, zygotes and embryos and to optimize the procedural efficiency of techniques such as intracytoplasmic sperm injection (ICSI), assisted hatching, and embryo biopsy [9,10,11].
For male gametes, in vitro manipulations include techniques for sperm selection, sorting, immobilization and other incubation procedures. The first use of lasers to manipulate human gametes can be traced to 1984 [12]. Sato et al. were the first to report the use of lasers to manipulate human sperm and explore the effects of laser exposure on sperm motility and velocity in vitro [12]. Over the next few years, near-infrared (NIR) laser beams with wavelengths ranging from 700 to 1200 nm were used as optical traps (laser tweezers) in sperm micromanipulation [13,14,15]. Further development of laser applications was the introduction of an infrared diode laser emitting at a wavelength of 1480 nm, which is far from the absorption peak of DNA (260 nm) [16]. The system allows laser beams along the microscope’s optical axis to the target with minimal absorption by the culture dish and the water molecules [9, 17]. This 1480 nm laser is used across a variety of applications, including sperm selection, sorting, immobilization prior to ICSI, and viability assessment of immotile sperm.
Many studies have been performed to investigate the different laser technologies in in vitro manipulations of human spermatozoa. However, there is a lack of a unified understanding of laser application in the manipulation of spermatozoa and, subsequently, the inability to make clear and accurate decisions on the clinical value of these laser technologies. The present review aims to summarize the advancements and improvements of laser technologies applied in the manipulation of human spermatozoa, such as photobiomodulation, sperm sorting, selection, and immobilization prior to ICSI. We also evaluate the potential value of these laser technologies in the treatment of male infertility and safety considerations for clinical application.
Photobiomodulation therapy on human spermatozoaPhotobiomodulation (PBM) therapy, previously termed low-level laser therapy (LLLT), generally employs light at red and NIR wavelengths to modulate biological activity [18]. Several studies were conducted in vitro on human spermatozoa through the employment of low-level laser therapy, demonstrating a positive effect on sperm function. PBM parameters have been mostly reported within the red and NIR wavelength range of 600–1100 nm, with an energy density of between 5 and 200 mW/cm2. PBM laser devices commonly include Krypton Laser, gallium aluminum arsenide (GaAlAs), neodymium-doped yttrium aluminum garnet, and indium gallium aluminum phosphide (InGaAlP) diode lasers. Table 1 presents a summary of the results.
Table 1 Effect of low-level laser therapy (LLLT) on human sperm parameters Effects of PBM on the parameters of human spermatozoaSperm motilitySperm motility is one of the most important characteristics associated with fertility. Almost all studies have examined the impact of PBM therapy on the motility and other kinematic parameters of human sperm. Sato et al. reported that normal sperm samples were exposed to red laser at different dosages, 0.5, 1.0, 2.0, 4.0, 8.0, and 32 J/cm2, for 160 s or 80 s and found that total sperm motility significantly increased at 4.0, 8.0 and 32 J/cm2 [12]. However, there was no stimulating effect on sperm velocity [12]. A study by Preece et al. assessed the effect of a 633 nm red laser at a power density of 5.66 mW/cm2 on frozen human sperm and indicated that the swimming speed improved within 35 min of irradiation [19]. In another work, human fresh and frozen sperm were exposed to light from a GaAlAs single laser beam (810 nm, 200 mW) and an LED cluster (660 and 850 nm, total power 2 W) for different irradiation times [20]. The results showed that the change in sperm motility was dependent upon the stimulatory dose, exposure time, and condition of the sample [20]. Concerning the effects of red and infrared laser irradiation on the motility of sperm, Safian et al. performed three different energy densities of two wavelengths of laser [21]. The results concluded that the NIR laser at 0.6 J/cm2 density was superior to the other irradiation protocols in stimulating the effect on motility [21]. The results from the study of Lenzi et al. indicated that the increase in progressive sperm motility after laser irradiation (647 nm) was related to the fast consumption of sperm ATP contents, suggesting that laser irradiation may have an “energetic modulation effect” on normal sperm [22]. However, one study reported by Highland et al. found that NIR irradiation resulted in a damaging effect on sperm viability and a diminished membrane function of sperm [23]. In addition, several studies have used laser beams to stimulate abnormal sperm, including sperm from oligospermia and asthenospermic patients, and showed that irradiation significantly increased sperm motility and velocity [24,25,26]. In two studies by Safian et al., PBM therapy before human sperm cryopreservation dramatically increased the percentage of live spermatozoa [27, 28].
DNA integritySperm DNA integrity is critical for the success of fertilization, embryo development, and implantation and is therefore considered a predictive factor for the clinical outcomes of patients undergoing ART. The potential effect of laser light on sperm DNA integrity has been a focus of attention by investigators. Most published studies have confirmed that red and infrared light does not induce DNA damage. Firestone et al. reported that infrared laser irradiation did not cause any increase in DNA damage at 2 h after exposure in normospermic, oligospermic, and asthenospermic samples [26]. Similarly, exposure to a pulsed-wave laser had no significant effect on the DNA fragmentation level in sperm from asthenozoospermic patients [24]. 830-nm laser irradiation can slightly increase the level of DNA fragmentation in sperm from asthenospermic patients, but it was not statistically significant [25]. The results from the study of Preece et al. showed that red light exposure could not produce sufficiently high levels of reactive oxygen species (ROS) to cause significant oxidative damage in normal sperm DNA [19]. Consistent with previous research, Gabel et al. also found no damage to sperm DNA integrity by light irradiation at a very high-density dose [20]. Interestingly, Safian et al. studied the effects of red and NIR ranges of PBM with a diode laser alone and together on the DNA fragmentation index (DFI) of fresh human sperm and observed that compared with the control, both the red + NIR and red lasers significantly increased DFI, while the NIR range of PBM did not result in a detectable increase in DNA damage [21].
Mechanism of PBMAlthough the underlying pathways of PBM therapy are not well established and may vary among different sperm states (fresh versus frozen, normal versus abnormal), laboratory and clinical studies suggest that PBM significantly improves sperm motility and does not damage DNA. It has become increasingly clear that the biological effects of PBM are closely associated with dosage and irradiation time as well as sperm condition. The lack of consistency in study conditions has always been a confounding factor in the interpretation of PBM mechanisms. Nonetheless, several studies have addressed probable mechanisms regarding the interaction of laser light and spermatozoa.
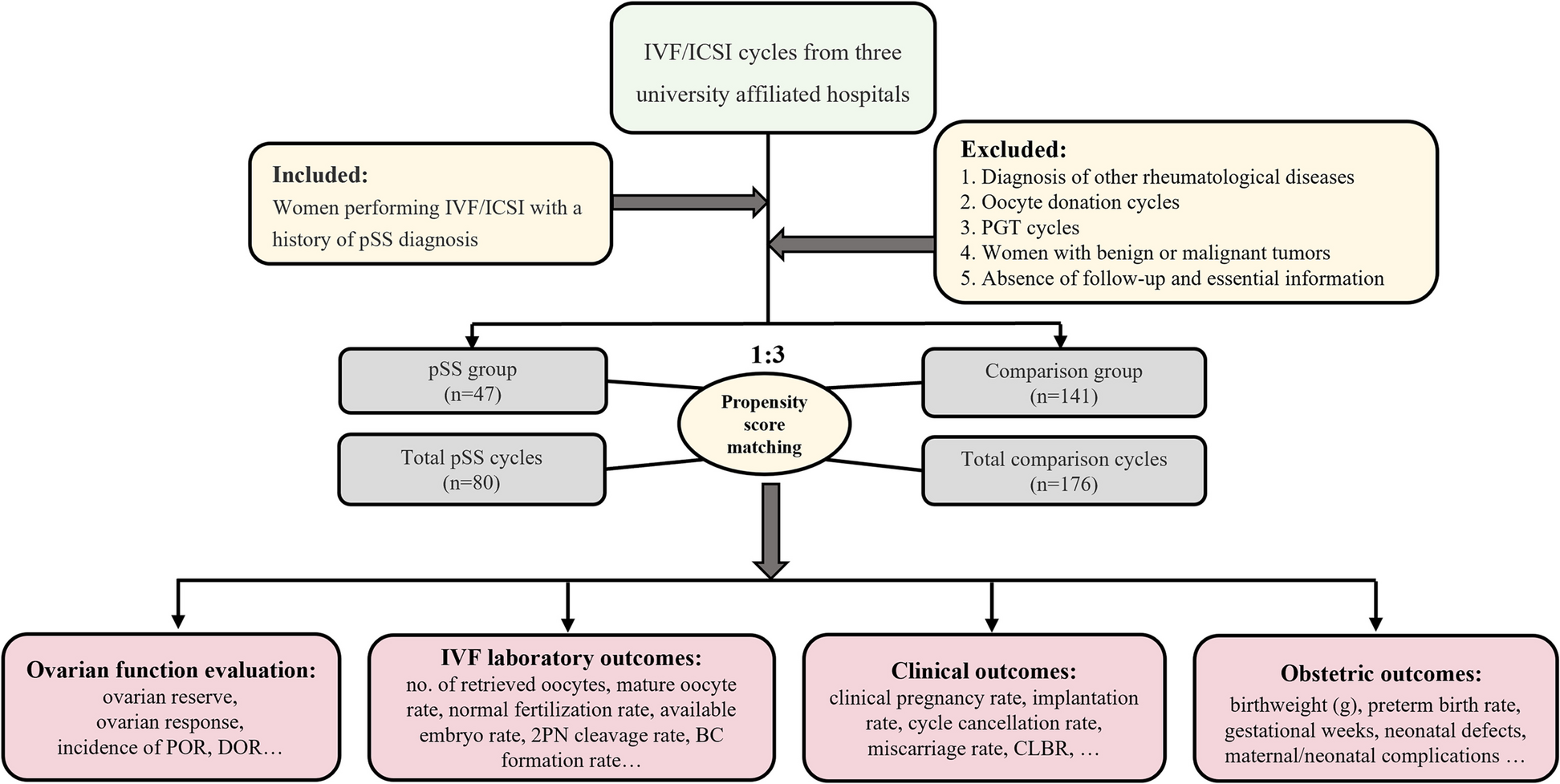
At present, at least four mechanisms are believed to be related to the response of sperm to PBM. The first hypothesis suggests that the effects of irradiation are driven by changes in mitochondrial function. Current data indicate that PBM mainly acts on cytochrome c oxidase (CcO) in the mitochondrial respiratory chain [29]. Photonic energy in red or NIR light is absorbed by CcO, resulting in an increased transmembrane proton gradient that drives the production of adenosine triphosphate (ATP), thus increasing sperm motility [19, 20, 30, 31]. ATP is the universal energy source in sperm cells essential for the maintenance of motility driven by flagellar dyneins. An increase in ATP synthesis leads to increased activity of all ATP-driven carriers for ions, such as Na+/K+ ATPase and Ca2+ pumps [32, 33]. Ca2+ is a key regulator of sperm motility. Furthermore, since ATP is the substrate of adenylate cyclase, ATP levels control the level of cAMP [34]. Both Ca2+ and cAMP are very important second messengers [35, 36]. The second mechanism considers the generation and/or release of reactive oxygen species (ROS) from sperm cells after laser light irradiation. ROS are very small molecules, including oxygen ions, free radicals, and hydrogen peroxide. Previous studies have shown that exposure of sperm to light increases hyperactivated motility mediated by mitochondrial ROS production and the cAMP/PKA pathway [37, 38]. Using the electron paramagnetic resonance (EPR) spin-trapping technique, Lavi et al. followed light-induced hydroxyl radicals in sperm cells and found that the concentration of hydroxyl radicals increased with illumination time and that ROS were produced in both the membrane and cytoplasm [39]. According to a study by Shahar et al., light-stimulated hyperactivated motility was increased through ROS-dependent activation of the epidermal growth factor receptor (EGFR) [38]. The third mechanism for PBM involves nitric oxide (NO), which is an important biological messenger that plays a crucial role in the regulation of energy production and mitochondrial biogenesis [40]. Previous studies revealed that light can induce NO formation by increasing the activity of nitric oxide synthase (NOS) [41,42,43]. NO is important in sperm motility and capacitation [44, 45]. The fourth hypothesis is linked to the interaction between light and specific receptors of the opsin family, which are coupled to G-proteins in sperm [46]. There is evidence that sperm opsins, specifically rhodopsin, play a role in the response of sperm to light [47]. At least seven opsin proteins are present in human sperm, of which the contents of encephalopsin and neuropsin are the most abundant [48]. The opsins, as light transducers, work through the light activation of photosensitive molecules linked to opsins [47]. The exact function of these opsins in the response of sperm to light still requires further study. Schematic diagrams of four possible mechanisms of PBM therapy on sperm are presented in Fig. 1.
Fig. 1
Diagram illustrating four possible mechanisms of PBM therapy on human sperm. Photonic energy in low level light is absorbed by the enzyme cytochrome c oxidase (CcO) located in the mitochondrial respiratory chain. The activated enzyme leads to a proton gradient. Consequently, the levels of reactive oxygen species (ROS) and adenosine triphosphate (ATP) are increased. On the other hand, the application of low level light activates specific receptors of the opsin family coupled to G-proteins in sperm. Phosphatidylinositol 4,5-diphosphate (PIP2) is catalyzed by hydrolysis to produce inositol 1,4,5-triphosphate (IP3). All of these activities activate light-sensitive ion channels and increase the levels of calcium ions (Ca2+). Soluble adenylyl cyclase (sAC) is activated by Ca2+. The increased sAC activity activates 3’,5’-cyclic adenosine monophosphate (cAMP)/ protein kinase A (PKA) pathway, thereby promoting sperm motility. In addition, exposure to low level light induces nitric oxide (NO) production by nitric oxide synthase (NOS), which activates cyclic guanosine monophosphate (cGMP)-dependent protein kinases production from soluble guanylate cyclase (sGC). Sperm motility is promoted by activation of protein kinase G (PKG).
PBM therapy for male infertilityPBM therapy is a fast-growing technology and provides a promising tool for improving male infertility status [49]. Currently, only a small number of studies have been carried out to evaluate the effect of PBM on the functional capacity of sperm from oligo- and astheno-zoospermia patients [24,25,26, 50]. Due to a variety of protocol parameters, it is difficult to compare directly between different studies. However, the results of these studies reveal a similar trend; PBM positively affects sperm motility and velocity without causing any damage to the DNA in samples of oligo- and astheno-zoospermia patients [24,25,26]. In addition, PBM therapy prior to human sperm cryopreservation plays a significant role in improving the quality of postthawed sperm and preventing cryo-damage [21, 28]. However, few studies have conducted assessments of human sperm function, such as acrosome reaction, hyperactivation, and fertilization ability. Recent studies evaluated the effect of PBM therapy on the improvement of spermatogenesis in hyperthermia-induced azoospermia mouse models and found that the spermatogenesis process is significantly improved by PBM therapy [51, 52]. In addition, accumulated evidence from animal studies suggested that PBM therapy improved sperm capacitation and fertilizing ability, as well as reproductive performance [53, 54]. It is postulated that PBM therapy can be considered a promising first-line medical intervention in the treatment of male infertility in the future [30, 31].
Given a lack of guidelines and a limited evidence base for PBM treatment on human sperm, the issues outlined below have been highlighted and addressed for consideration. First, the mechanisms responsible for the beneficial effect on sperm reported by PBM therapy need to be completely elucidated. Theoretically, sperm cells are not exposed to any type of light, so it can be speculated that both normal and abnormal sperm can be sensitive to exogenous bright stimuli. Although different hypotheses have been proposed to explain the effects of light irradiation on sperm, future research is needed to clarify which mechanism plays an important role. Further understanding of the mechanisms is necessary for optimizing clinical treatment. Second, PBM treatment protocols need to be optimized for each type of sperm, such as normal sperm and sperm with mild, moderate, and severe asthenozoospermia. There is now a wide and increasing array of laser equipment to choose from. The effectiveness of PBM therapy is likely to depend on specific laser characteristics. To achieve a desirable clinical outcome, the proper wavelength, pulse duration and energy density must be tailored to the clinical indication. Finally, to evaluate the suitability of PBM for routine clinical use, it is necessary to further study whether PBM has potential genotoxic effects on sperm. DNA integrity is considered a fundamental factor for the fertilization and transmission of paternal genetic information to offspring. Further research on the effects of low-level lasers on sperm cells is imperative.
Taken together, several general conclusions can be drawn: (i) The mechanical basis of PBM therapy is associated with several intracellular metabolism pathways that regulate sperm motility. (ii) The overall results from preclinical and clinical studies suggest that PBM therapy holds promise as a non-invasive treatment for male infertility disorders, such as asthenospermia and oligospermia, by enhancing sperm motility and quality. (iii) PBM beneficial effects depend on wavelengths, exposure time, stimulatory dose, irradiated area, and other treatment parameters (e.g., condition of the sample). Tailoring PBM treatment protocols for specific sperm conditions is essential.
Micromanipulation of human sperm using a laser optical trapAn optical trap is a noninvasive biophotonic tool that has been studied for practical applications in a variety of fields, including physics, chemistry, biology, and medical science [55, 56]. A laser optical trap was first reported to manipulate single cells in 1987 [57]. The authors achieved damage-free trapping and manipulation of suspensions of single cells and organelles located within individual living cells [57]. Berns et al. first showed that a laser trap could be used to move the chromosomes inside mitotic cells in vitro [58]. Not long afterward, single-point laser traps were used as a tool to manipulate individual sperm cells and analyze the interaction between laser and sperm and sperm motility by measuring sperm swimming forces in the late 1980s [13, 59]. Several studies have demonstrated that laser optical trapping and micromanipulation of sperm cells using a NIR beam is technically feasible [14, 15, 60,61,62,63,64,65,66,67,68,69,70].
Laser trapping optical systemThe optical trapping system is designed as a biomedical tool to study the physiological and biomechanical properties of cells. This system introduces NIR laser light into an inverted microscope, creating a single-point and three-dimensional gradient laser trap at the focal point of the microscope. Trapping provides a noninvasive method for analyzing and classifying sperm based on sperm swimming speed and swimming force [13,14,15, 60, 65,66,
留言 (0)