

記住我



Until recently, the cerebellum was regarded as a brain structure engaged mainly in motor control. Studies dating back to the early 19th century showed that cerebellar damage in pigeons resulted in an inability to fly due to uncoordinated wing movements (Flourens, 1824). After the cerebellum’s role in motor control was confirmed in studies on monkeys, it became a fundamental truth in clinical neurology and brain sciences (Russell and Horsley, 1894). Since then, numerous studies from the 20th and 21st centuries have reaffirmed the cerebellum as a structure that plays a vital role in motor control and motor learning (Manto et al., 2012). However, though the fields of cognitive neuroscience and experimental neuropsychology did not emerge until the early 20th century, other researchers in the 19th century reported emotional and intellectual impairment in patients with cerebellar agenesis (Fisher, 1839). Yet, it wasn’t until the 1990s that researchers refocused on the cerebellum’s non-motor functions (Schmahmann, 1991). Studies on cerebellar patients revealed significant impairments in executive function and attention (Stoodley and Schmahmann, 2010). Cerebellar abnormalities also have effects on other neurocognitive domains. These include language, visuospatial perception, visual construction ability, episodic memory, and social cognition (Hoche et al., 2016; Mariën and Borgatti, 2018; Slapik et al., 2019; Ray et al., 2020).
In line with these clinical findings, neuroimaging studies reveal that the cerebellum is anatomically and functionally interconnected with areas of the brain that are important for cognitive and emotional functions (Stoodley and Schmahmann, 2010). There is an abundance of mutual connections between the cerebellum and other areas of the brain, including the prefrontal cortex, the posterior parietal cortex, and the basal ganglia (Middleton and Strick, 1994; Kelly and Strick, 2003; Bostan et al., 2013).
This can explain why disorders affecting the cerebellum also lead to disrupted affective and cognitive functions, such as the cerebellar cognitive affective syndrome (CCAS) (Schmahmann and Sherman, 1998; Stoodley and Schmahmann, 2010; Ahmadian et al., 2019). Moreover, the profile and severity of CCAS symptoms varies in a predictable pattern depending on the pathological process and what part of the cerebellum is affected (Argyropoulos et al., 2020).
Here, we review literature on the anatomy and physiology of the cerebellum, and the neurocognitive functions of three conditions that have been linked to cerebellar function: SCA3, ADHD, and ASD. SCA3 is a neurological disease where cerebellar degeneration is clearly and robustly demonstrated, ADHD and ASD are neuropsychiatric disorders that have been associated with cerebellar abnormalities, but the extent of cerebellar involvement is not as pronounced as in the case of SCA3 (Riva et al., 2013; Wan et al., 2020; Duan et al., 2021; Stezin et al., 2021). We selected these three conditions to highlight the cerebellum’s role in cognition and how cognitive effects vary according to the severity of cerebellar abnormalities, more evident in SCA3 and lighter in ADHD and ASD.
We attempt to link these findings to contemporary theories and models of the cerebellum, and we discuss how symptoms may arise from interactions between the cerebellum and other brain parts that influence cognitive functions. We then discuss potential clinical implications from knowledge about the cerebellum’s involvement in cognitive functions. A cerebellar-dependent neuropsychological test battery could be a valuable tool in the diagnostic process when data from medical examinations such as biomarkers and neuroimaging data are missing or are unclear. This could lead to an improved understanding of the neurocognitive functions of these patient groups, and, ultimately, better treatment.
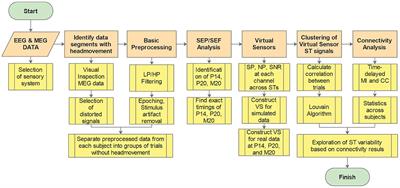
Methods Search strategy and study selectionWe performed a literature search in two electronic databases: PubMed and Embase. Through these searches, we identified articles supplying information on the neurocognitive profile, neurocognitive functions, and cerebellar functions of patients with three specific diagnoses ADHD, ASD, and SCA3.
Original articles and chapters reporting relevant studies on humans and animals were included. We used terms such as “ADHD,” “ASD,” and “SCA3” coupled with terms related to the cerebellum, cerebellar functions and to neurocognitive domains and neurocognitive profiles, such as “cognition,” “intelligence,” “visual perception/construction,” “processing speed,” “sensorimotor integration,” “executive function,” and “memory.”
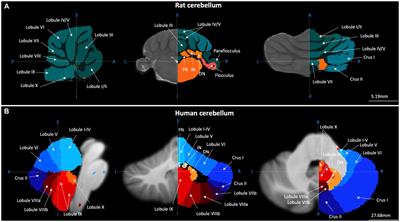
CerebellumThe cerebellum (literally “little brain” in Latin) contains 80,2% of all neurons in the human brain as well as 19% of all the non-neural cells in the brain (Azevedo et al., 2009). The anterior lobe of the cerebellum receives afferents from the spinal cord through the spinocerebellar tracts and from the cerebral cortex via corticopontine projections (Brodal, 1978; Hartmann-von Monakow et al., 1981; Schmahmann et al., 2004). The posterior lobe receives input mainly from the brainstem and cerebral cortex (Siegel and Sapru, 2006). The cerebellar vermis and fastigial nucleus are interconnected with the vestibular nuclei and other brainstem nuclei engaged in motor control, gait, and equilibrium, and with brainstem nuclei linked with limbic and paralimbic cortical and subcortical regions (Voogd, 2004).
The cerebellum is not only connected with structures concerned with motor control. Large parts of the cerebellum forms reciprocal connections with association areas of the cerebral cortex including the prefrontal cortex, posterior parietal cortex, superior temporal polymodal regions, cingulate gyrus, and posterior parahippocampal area (Strick et al., 2009; Bostan et al., 2013). The anatomical circuits linking the cerebellum with the cerebral cortex are organized in a two-stage feed-forward loop (the corticopontine and pontocerebellar projection) and a two-stage feedback loop (cerebellothalamic-thalamocortical projection). These loops are constituted by multiple parallel but partially overlapping subcircuits that link different areas of the cerebral cortex with specific regions of the cerebellum (Brodal, 1978; Voogd and Glickstein, 1998; Voogd, 2004; Apps and Garwicz, 2005). The cerebellum is also mutually interconnected with the basal ganglia via oligosynaptic loops (Hintzen et al., 2018). These anatomical connections hint that the cerebellum plays a role that goes beyond motor control. Indeed, as we will discuss below, evidence from recent decades suggests that the cerebellum is highly involved in a wide spectrum of cognitive functions.
Output from the cerebellar cortex goes via the cerebellar nuclei. The medial (fastigial) nucleus projects mainly to the brainstem and spinal cord; the interposed (comprising the emboliform and globose) nucleus targets mainly the midbrain and the thalamus; and the lateral (dentate) nucleus projects mainly to the thalamus. Some neurons in the cerebellar nuclei project directly to the red nucleus, which then projects to other nuclei that control muscles. This is how the cerebellum can elicit blink responses even in the absence of the cortex (Hesslow and Yeo, 2002). More recently, it was discovered that the cerebellar nuclei also contain neurons that project to the Purkinje cells forming the nucleo-cortical cerebellar pathway (Ankri et al., 2015). Apart from the inhibitory input from the Purkinje cells, the cerebellar nuclei also receive excitatory input from collaterals of both mossy fibers and climbing fibers (Ten Brinke et al., 2017).
Functions of the cerebellumKnowledge of cerebellar functions has been derived mostly from studying natural and experimental ablative lesions (Fine et al., 2002). The cerebellum plays a crucial role in making predictions and creating feedforward models that guide our behavior (Wolpert et al., 1998; Welniarz et al., 2021). Consequently, cerebellar damage makes movements jerky, slow, and uncoordinated–as is evident in ataxia and dystonia (Therrien and Bastian, 2015). In line with the cerebellum’s role in motor coordination, disruption of the ionic homeostasis in cerebellar neurons has been shown to evoke motor impairments in mouse model studies (Hoxha et al., 2018). The impairments range from gross abnormalities to more specific impairments. For example, disruption of the high-voltage activated Ca2+-influx in Purkinje cells severely disrupts action potential firing and motor behavior in mouse model studies (Tara et al., 2018). Likewise, deletion of several chloride channels and transporters in mouse models has been associated with motor abnormalities (Poroca et al., 2017). For instance, knockout mouse models with deletion of GABAA receptors from Purkinje cells did not show ataxia, but they displayed significant impairments in several aspects of locomotion, motor performance and motor learning (Wulff et al., 2009).
Clinical research from the last three decades show that the cerebellum is not merely concerned with motor functions. The human cerebellum is linked with cerebral association areas concerned with intellect, social cognition, perception, recognition, forwarding and encoding of emotional information, and the regulation of emotional states in relation to cognitive, motor and social behaviors (Stoodley and Schmahmann, 2009a,b, Stoodley et al., 2010; Adamaszek et al., 2017). In line with the anatomical connections between the cerebellum, the basal ganglia (K B et al., 2022), and the cerebral cortex (Allen and Tsukahara, 1974; Holdefer et al., 2000; Strick et al., 2009), the cerebellum has been shown to play a role in many cognitive and emotional functions (Reeber et al., 2013). The limbic/cognitive cerebellum is identified in the cerebellar posterior lobe; but also, others areas of the cerebellum contribute to cognition like the sensorimotor cerebellum represented in the anterior lobe (Schmahmann, 2019). Furthermore, cerebellar damage is associated with lesions in the posterior lobe resulting in deficits in executive functions, visuospatial processing, language abilities and affect regulation (Karatekin et al., 2000; Grill et al., 2004; Manes et al., 2009). Other disorders linked to morphological abnormalities in the cerebellum, such as ASD, schizophrenia, and ADHD have also been linked with affective dysregulation (Bernard and Mittal, 2014; D’Mello et al., 2015; Stoodley, 2016; Kim et al., 2021). The cerebellum also contributes to visual cognition. Patients with degenerative cerebellar disorders showed impairments on mental rotation, an operation that requires continuous manipulation of a visual representation (McDougle et al., 2022). Further, the cerebellum is involved in face and emotion processing via its connections with limbic and cortical areas important for processing facial emotions perception (Fusar-Poli et al., 2009; Schutter et al., 2009). The cerebellum is involved as well in timing (Ivry and Keele, 1989).
Neuroimaging, cognitive and behavioral findingsNeuroimaging studies have verified that the cerebellum contributes to a broad range of cognitive functions, like perception, language, working memory, cognitive control, and social cognition (Diedrichsen et al., 2019). A review of clinical, experimental and neuroimaging studies shows that sensorimotor tasks have been linked to the anterior lobe, while higher function tasks, including language, executive function, and working memory, have been linked to the posterior lobe of the cerebellum. Processing of emotions was localized to vermal lobule VII and lateral posterior hemisphere (Stoodley and Schmahmann, 2009a). The involvement of the cerebellum in these processes is due to functional connections with frontal lobes via the cerebello-thalamo-cortical and cortico-ponto-cerebellar loops (Middleton and Strick, 1994; Kelly and Strick, 2003).
Functional magnetic resonance imaging (fMRI) shows cerebellar activation during many cognitive, affectional, and social tasks in healthy participants, although there is significant inter-participant variability in the pattern of topographical activation on a specific task (King et al., 2019). A meta-analysis concluded that specific parts of the cerebellum were activated during cognitive tasks and music elaboration/processing (E et al., 2014). One of the first positron emission tomography (PET) studies examining the cerebellum by Petersen et al. (1988) showed activation in the right lateral cerebellum when participants generated words. The cerebellum is also involved in lexical retrieval, syntax, aphasia, reading, writing and metalinguistic skills (De Smet et al., 2013). Other cognitive functions like working memory, social cognition, emotional memory, semantic memory and visuospatial abilities are also linked to the cerebellum (Ashida et al., 2019; Slapik et al., 2019; Van Overwalle et al., 2020; Gatti et al., 2021; Fastenrath et al., 2022). Another fMRI study showed that the dentate, the output nucleus of the cerebellum, was activated by cognitive processing, for example, during a realization of a puzzle that required multiple cognitive processes (Kim et al., 1994).
Petacchi et al. (2005) analyzed studies that used fMRI and PET techniques, their analysis shows that specific regions in the cerebellum are activated during auditory tasks and contribute to sensory auditory processing. Recent results demonstrated that the cerebellum shows increased activity during enhanced emotional memory encoding; the midline cerebellum seems to be activated during recall of emotional personal life episodes, remarking an aspect of this region in emotional memory retrieval (Fastenrath et al., 2022).
Neuroimaging studies have shown that explicit motor timing activates the cerebellum, the supplementary motor area, basal ganglia, right-inferior frontal, and parietal cortices (Avanzino et al., 2016). Timing ability affects sequential rhythmic movements, motor control, and many cognitive tasks used to assess executive functions (Homack et al., 2005). The cerebellum is essential in timing-dependent learning processes. It plays a role in finger tapping, in which the participant must maintain a rhythm, and eyeblink conditioning, a simple form of associative motor learning in which subjects learn to associate a neutral stimulus with a reflex-eliciting stimulus (Gerwig et al., 2007; Bracha et al., 2009). The cerebellum, together with other brain areas like the primary sensorimotor cortex, the supplementary motor area, and the occipital cortices, are necessary to complete the finger-tapping task in response to visual cues (Turesky et al., 2018). Previous studies have brought to light that the main neuronal plasticity responsible for generating the conditioned eyeblink response takes place in specific regions of the cerebellum (Steinmetz, 2000; Thürling et al., 2015).
Spinocerebellar ataxia type 3 (SCA3)Ataxia derives from Greek and means “lack of order.” SCA is a rare inherited (autosomal dominant), progressive, neurodegenerative, and heterogeneous disease mainly affecting the cerebellum. SCA is the used nomenclature for Spinocerebellar Ataxia from type 1 to type 48 for autosomal dominant cerebellar ataxia a (Kuo, 2019). SCA3 is the world’s most common, dominantly inherited ataxia. Ataxia is characterized by decreased voluntary muscle coordination and impairment of movement control that affects gait stability, speech, and eye movement. Ataxia refers to the pattern of impaired movement and the group of diseases characterized by this specific symptom. The primary manifestation is losing balance while standing and walking (Ashizawa and Xia, 2016). Ataxia can be inherited, acquired, or idiopathic. Based on the inheritance mode, cerebellar ataxias are classified as Autosomal Dominant Cerebellar Ataxias, also known as SCA, Autosomal Recessive Cerebellar Ataxias, mitochondrial or X-linked ataxias (Durr, 2010; Klockgether, 2010). Acquired ataxias can be caused by many factors, such as strokes, tumors, multiple sclerosis (Kuo, 2019), alcoholism, peripheral neuropathy, metabolic disorders, or vitamin deficiencies (Klockgether, 2010). Cerebellar lesions associated with ataxia are often seen in neuroimaging (Kuo, 2019).
Due to the effects on the cerebellum, CCAS is present in SCA3 (Paulson, 2012). However, CCAS has been studied in SCA3 (Braga-Neto et al., 2012; Maas et al., 2021). Dystonia, depression, diplopia, sensorimotor polyneuropathy, restless legs syndrome, and parkinsonism are the recognized clinical symptoms of SCA3 patients (Kuo, 2019). Several genetic causes exist for ataxia; the CAG-repeat expansion is the leading cause (Durr, 2010). Besides the effect on the cerebellum (see below), the connectivity is impaired in both the striatal-cortical and cerebellar-cerebral networks (Lindsay and Storey, 2017). The striatal-cortical network, exactly the prefrontal striatal circuit, sustains a decisive role in higher-order cognitive functions (Alexander et al., 1986).
Cerebellum and ataxiaSCA3 influences the brain stem, cerebellum, and pyramidal tracts (D’Abreu et al., 2012; Wu et al., 2017). However, other studies also show an effect on other brain regions, such as basal ganglia, pallidum, and diencephalon (Arruda et al., 2020). Morphometric MRI has demonstrated typical patterns of atrophy or volume loss in the cerebellum and brainstem with ulterior lesions in some supratentorial areas (Wan et al., 2020; Stezin et al., 2021; van der Horn et al., 2022; Yap et al., 2022a). Some specific focal or regional atrophies noted in certain SCAs are present in SCA3, where studies show pontocerebellar atrophy with enlargement of the fourth ventricle (Bhandari et al., 2022). Animal models helped to clarify the pathogenesis of the cerebellar ataxias (Burright et al., 1995; Serra et al., 2006). Purkinje cell degeneration is one of the primary mechanisms in animal models of ataxia (Chen et al., 2008; Liu et al., 2009). Data from calcium imaging and electrophysiological studies demonstrate that Purkinje cells and their significant inputs are malfunctioning before the onset of degeneration in ataxia mouse models (Barnes et al., 2011; Hourez et al., 2011; Shakkottai et al., 2011; Kasumu et al., 2012; Hansen et al., 2013). Alterations of cerebellar function can be identified with motor learning paradigms that depend on the integrity of the olivocerebellar circuit, such as eyeblink. A recent study shows that this type of learning is impaired in preclinical carriers of a SCA3 mutation (van Gaalen et al., 2019).
The cerebellum has enduring subcortical and cortical connectivity, resulting in non-motor symptoms that are often neglected during clinical assessments in many neurodegenerative conditions, especially if clinical ataxia is not present (Schmahmann et al., 2004; Moro et al., 2019; Fastenrath et al., 2022). More than forty SCA have been recognized, SCA does not affect only the cerebellum, and SCA3 may involve other areas of the central nervous system, like pontine nuclei, spinal cord, cortex, peripheral nerves, and basal ganglia (Bhandari et al., 2022). Neural functions can be altered even if there are no visible imaging results, and these can be the primary cause of behavioral symptoms in cerebellar ataxia. Furthermore, cerebellar gray matter atrophy is found in several other neurodegenerative diseases such as amyotrophic lateral sclerosis, Alzheimer’s disease, frontotemporal dementia, Huntington’s disease, multiple systemic atrophy, Parkinson’s disease, and progressive supranuclear palsy (Gellersen et al., 2017). These studies together highlight the cerebellum’s role in both motor and cognitive functions, and they suggest that SCA affects motor function and cognition.
Neurocognitive functions in spinocerebellar ataxia type 3Although previously thought to primarily impact motor functions, numerous studies that we review below highlight that SCA3 also significantly impacts multiple cognitive domains.
Executive functionsPrevious studies show that SCA3 patients exhibit deficits on the Hayling Sentence Completion Test and the Stroop Color-Word Test–designed to assess inhibitory ability (Zawacki et al., 2002; Garrard et al., 2008; Moriarty et al., 2016). Feng et al. (2014) showed that the time required to complete Trail Making Test A did not differ significantly between SCA3 patients and controls, but it took significantly more time for SCA3 patients to finish Trail Making Test B. Results from Braga-Neto et al. (2012) showed that both Trail-Making Tests A and B were significantly more impaired in patients with SCA3 than controls. Patients with SCA3 have some deficits in auditory working memory, as measured by the Backward subtest of the Digit Span Test (Feng et al., 2014; Ma et al., 2014; Tamura et al., 2018). Several studies did, however, show average performance on Digit Span Test Forward, a measure of short-term memory that is more related to attention than to working memory (Zawacki et al., 2002; Bürk et al., 2003; Kawai et al., 2004; Braga-Neto et al., 2012; Lopes et al., 2013; Feng et al., 2014). Deficits in visuospatial working memory were also detected using Spatial Span and Corsi-Block tapping test (Braga-Neto et al., 2012; Lopes et al., 2013). The executive function impairments may demonstrate an interrupted cortico-striatal and cerebro-cerebellar network in patients affected by SCA3, coherent with similar cognitive difficulties highlighted in CCAS (Stoodley and Schmahmann, 2010; Lindsay and Storey, 2017).
Attention and processing speedContrasting results emerged for this cognitive domain. Yap et al. (2022b) described no impairment, while other studies show that SCA is associated with impaired visuospatial attention and processing speed (Zawacki et al., 2002; Braga-Neto et al., 2012; Tamura et al., 2018). The grade of ataxia severity, visuomotor search and cerebellar oculomotor problems may interfere with performance on neurocognitive tests (Braga-Neto et al., 2012; Tamura et al., 2018). On the other hand, auditory attention is not impaired in SCA3 (Garrard et al., 2008). The Symbol Digit Modalities Test and Elevator Counting subtest of the Test of Everyday Attention show impairment related to attention and process speed (Moriarty et al., 2016).
LanguageGiven that the cerebellum is also involved in language, it is perhaps unsurprising that SCA is associated with language problems (Mariën and Borgatti, 2018). One study on twenty SCA3 patients demonstrated that semantic fluency, phonemic fluency, and category switching were significantly lower in SCA3 patients compared to healthy controls (Maas et al., 2021); the same results were found in a longitudinal study (Moriarty et al., 2016). Verbal fluency was assessed in letter and category fluency tasks, it was also found to be impaired in multiple studies (Kawai et al., 2004; Klinke et al., 2010; Lopes et al., 2013; Feng et al., 2014; Ma et al., 2014; Tamura et al., 2018).
Besides, contrasting test results emerge for semantic verbal fluency with and without impairment (Bürk et al., 2003; Kawai et al., 2004; Garrard et al., 2008; Klinke et al., 2010; Braga-Neto et al., 2012; Feng et al., 2014; Tamura et al., 2018). According to some studies, the naming ability of SCA3 patients, an aspect of the semantic memory which requires retrieval capacity, is not impaired (Zawacki et al., 2002; Kawai et al., 2004; Lopes et al., 2013).
To some extent, impairment in executive functions may mediate the language deficits seen in patients with SCA. For example, patients may have trouble with word retrieval that implicates the cerebellum and its connections with the cerebrum (Mariën et al., 2014). Fundamental executive dysfunction in patients with SCA3 may interfere with the retrieval processes of verbal information from memory storage (Tamura et al., 2018). Divergent phonemic and semantic verbal fluency results indicate the cerebellum’s role in the motor and sequential speech mechanisms. Cerebellar impairment can affect verbal fluency by explicitly affecting the performance of phonemic rules. These results highlight the importance of computational properties in developing linguistic strategies by the cerebellum (Leggio et al., 2000).
MemoryPatients with SCA also exhibit memory problems. Several studies show that SCA3 patients have deficits with immediate auditory recall and delayed auditory recall (Bürk et al., 2003; Kawai et al., 2004; Garrard et al., 2008; Lopes et al., 2013; Feng et al., 2014; Ma et al., 2014; Tamura et al., 2018). A single study shows impairment of delayed recall but average immediate recall (Elyoseph et al., 2020). Deficits in both immediate and delayed recall in visual episodic memory were identified with the Visual Paired Associate test, and delayed recall in visual episodic memory was detected with the Visual Reproduction test. Scores were independent of ataxia severity even though motor coordination is necessary to complete this test (Kawai et al., 2004). Different results were obtained in another study that utilized the Visual Reproduction Test of Wechsler Memory Scale-III, where immediate visual recall was poorer while delayed visual recall was intact (Garrard et al., 2008).
SCA3 patients did not show any visual recall impairment measured with the Rey-Osterrieth Complex Figure test, a non-process-pure test that can measure more cognitive functions (Meyers and Kr, 1995; Bürk et al., 2003; Braga-Neto et al., 2012). Mainly, auditory and visual recognition memory was intact in several studies (Zawacki et al., 2002; Garrard et al., 2008; Klinke et al., 2010; Lopes et al., 2013; Elyoseph et al., 2020). A study that used the Recognition Memory Test showed that SCA3 patients have more impairment in face recognition than word recognition (Moriarty et al., 2016). Moreover, auditory and visual recognition is usually unaffected, indicating that encoding and consolidation work and memory difficulties result from impaired free recall. Problems with recall are due to impairments of executive functions. A study shows contrasting results for procedural learning, where SCA3 patients have impaired procedural learning of the sensory-motor sequence on the procedural Serial Reaction Time Test but regular learning on the procedural Weather Prediction Probabilistic Classification test (Elyoseph et al., 2020).
The results on sequence learning impairment are coherent with the present opinion on dysregulated striatal-cortical and cerebellar-cerebral networks in SCA3 (Lindsay and Storey, 2017). Parietal and prefrontal areas are activated during mindful learning and recall of sequence information mediated by working memory, where the cerebellum and motor-related areas are accountable for the habituation process (Eliassen et al., 2001). An impaired auditory and visual recall is present in SCA3 patients. Recognition memory is intact; cognitive impairments influence information retrieval, which is necessary for free recall test conditions. These study results do not prove a temporal lobe impairment (Lalonde and Botez-Marquard, 2000), where recall and recognition memory are significantly impaired. However, these results could confirm the involvement of a dysregulated prefrontal-striatal network (Lindsay and Storey, 2017). These results are coherent with the cognitive profile of the cerebellar cognitive affective syndrome, where the memory is altered as a consequence of impaired executive functions (Ray et al., 2020). In conclusion, several studies show that SCA3 is associated with memory deficits–especially for memory functions related to the frontal cortex.
Visuospatial perception and visuospatial abilityIn contrast to the cognitive domains discussed so far, where there are evident deficits, at least in some sub-domains, the evidence for deficits in visuospatial perception and ability is relatively weak. Visual perception has been measured in SCA3 patients with the Incomplete Letters subtest of the Visual Object and Space Perception Battery. Results showed performances within the normal range (Garrard et al., 2008). No impairment was detected in studies that utilized the copy trial of the Rey–Osterrieth Complex Figure test, which measures visual-constructional ability and visual memory (Bürk et al., 2003; Braga-Neto et al., 2012). In contrast, the cube drawing test, which requires a three-dimensional perspective, showed impairments in this category of patients (Maas et al., 2021).
Social cognitionTheory of Mind is impaired in SCA3 patients (Moriarty et al., 2016). However, the ability to identify normative situations or violations of normative situations and attribute emotions like happiness, sadness, fear, anger or embarrassment is not impaired (Garrard et al., 2008; Sokolovsky et al., 2010). Difficulties emerged with the ability to describe, interpret, and justify the main character’s behavior during real social situations, presented in 14 stories. Social cognition has, however, rarely been examined in these patients, perhaps because SCA has always been treated as a disease that affects motor performance above other things. More research on the effects of cerebellar damage on social cognition is needed.
General intelligenceSeveral studies have investigated the impact of SCA3 on general intelligence, with contradictory results. A recent review by Yap et al. (2022b) concluded that general intelligence and neurocognitive functions such as verbal reasoning, semantic language, recognition, visuospatial perception, visuoconstructive ability, attention, and processing speed are relatively preserved in SCA3 patients (Yap et al., 2022b). Some studies reported the preservation of general intelligence, as measured by a composite score that includes both non-verbal and verbal reasoning (Zawacki et al., 2002; Bürk et al., 2003; Kawai et al., 2004; Kamphaus, 2005; Klinke et al., 2010), while one study underlined lower intelligence level compare to the control group (Garrard et al., 2008). However, one study reported lower general intelligence scores in SCA3 patients compared to the average population (Garrard et al., 2008). On the other hand, verbal reasoning appeared to be preserved, with average scores found on the Verbal Comprehension subtest of the Wechsler Adult Intelligence Scale (Zawacki et al., 2002; Bürk et al., 2003; Garrard et al., 2008; Klinke et al., 2010; Braga-Neto et al., 2012; Lopes et al., 2013). The results for non-verbal reasoning were less consistent with some studies demonstrating apparent impairment on the Perceptual Reasoning subtest of the WAIS (Kawai et al., 2004; Garrard et al., 2008; Braga-Neto et al., 2012), while others did not find any significant impairment (Zawacki et al., 2002; Bürk et al., 2003; Lopes et al., 2013). Overall, the literature suggests that general intelligence is, at most, only mildly impacted in SCA3 patients.
SummaryIn conclusion, multiple studies on SCA3 patients with cerebellar damage have shown impairments in various cognitive domains, particularly in executive functions, and secondary effects in other areas. The neurocognitive functions are illustrated in Table 1. The cognitive profile of SCA3 shows impairment in the executive functions of inhibition, cognitive flexibility, and visual and verbal working memory (Zawacki et al., 2002; Braga-Neto et al., 2012; Feng et al., 2014).
Table 1. Summarizes the evidence in the papers reviewed.
Previous results showed a link between activity in the cerebro-ponto-cerebello-thalamo-cortical loop and executive functions in SCAs (Lindsay and Storey, 2017). This is consistent with the previously reported core impairment of CCAS (Stoodley and Schmahmann, 2010; Lindsay and Storey, 2017).
Executive dysfunctions also affect verbal functions like semantic and phonemic fluency as well as fluency while switching between categories (Kawai et al., 2004; Moriarty et al., 2016; Maas et al., 2021). These are all functions that demand retrieval from long-term semantic memory. Semantic memory seems to be unaffected in these patients (Zawacki et al., 2002; Kawai et al., 2004; Lopes et al., 2013). Executive dysfunctions also impact episodic memory, with results showing evident deficits in information retrieval in visual and verbal episodic memory, while recognition and consolidation of information are partially intact (Bürk et al., 2003; Garrard et al., 2008; Braga-Neto et al., 2012; Moriarty et al., 2016). Visuospatial attention, visuospatial ability, and processing speed are also impaired (Zawacki et al., 2002; Moriarty et al., 2016; Tamura et al., 2018).
While social cognition is not well-studied in patients with SCA, some findings suggest that the Theory of Mind may be affected. Psychiatric conditions such as anxiety and depression, common among SCA3 patients, may also harm cognition. Language and cultural differences may also confound psychological studies, including those we have reviewed here. The severity of ataxia, the duration of the disease, and visual, articulatory, or upper limb motor difficulties can also influence cognition and hence the performance on neuropsychological assessments. However, because of the small sample sizes of previous studies, different confounders, and different neuropsychological methods that have been used, the neurocognitive profile of SCA3 needs to be better characterized and needs to be investigated further.
Attention deficit hyperactivity disorderADHD is a neuropsychiatric disorder present in approximately 5% of all children and in approximately 2.5% of the adult population (Regier et al., 2013; Austerman, 2015). It is estimated that about 15% of patients in adult psychiatry have ADHD (Deberdt et al., 2015). ADHD is defined based on clinical symptoms indicating dysregulation of attention and or activity/impulse control. When ADHD is suspected in adulthood, the investigation can be supplemented with the Wender Utah Rating Scale and Adult ADHD Self-Report Scale (Anbarasan et al., 2020).
Adults with ADHD show emotional dysregulation and executive function deficits (Barkley, 2010; Kessler et al., 2010; Biederman et al., 2019). While adults with ADHD often find ways to deal with attention problems and hyperactivity, the demands on executive abilities increase. Most jobs require complex planning and organizational skills, causing difficulties for adults with ADHD. Organizational problems often cause frustration and despair that lead to anxiety, sleep disorders, fatigue, depression or other conditions (Brattberg, 2006; Dirks et al., 2017; Schiweck et al., 2021). ADHD symptoms are normally distributed in the population, meaning they are present to varying degrees in many people. It is only when the symptoms are prominent and create a significant functional impairment that an ADHD diagnosis is justified. To receive the diagnosis, manifestations of the main symptoms of inattention and/or hyperactivity and impulsivity, along with clear functional problems due to ADHD symptoms, should be present before the age of twelve (Doernberg and Hollander, 2016). Patients with ADHD do not just look for treatment and assistance for what is classically perceived as ADHD, but for various psychiatric symptoms and functional problems, such as depression, fluctuating mood, fatigue, and anxiety (Brattberg, 2006). In summary, to diagnose ADHD is still challenging. This heterogeneous neuropsychiatric condition has many comorbidities with similar phenotypes like anxiety disorders, mood disorders, bipolar disorder, hypomania, substance use disorders, sleep disorders, Tourette syndrome, ASD and other behavioral disorders (Hegerl et al., 2010; El Malhany et al., 2015; Doernberg and Hollander, 2016; Fadeuilhe et al., 2021; Schiweck et al., 2021). Even if ADHD is considered a neurodevelopmental disorder, ADHD symptoms usually persist in late adulthood and is often confounded with mild cognitive impairment (Callahan et al., 2017).
Cerebellum and ADHDThe brain of patients with ADHD has been the subject of extensive research. One of the most robust findings is the abnormal structure and function of the cerebellum in patients diagnosed with ADHD. Abnormal cerebellar development has been implicated in ADHD (Vloet et al., 2006; Vaidya, 2011; Stoodley, 2016; Bruchhage et al., 2018). Functional imaging studies point to abnormalities in the cerebellum and other brain structures. Cerebellar and cerebral volume is ∼4% smaller in ADHD patients compared to controls (Castellanos et al., 2002; Vaidya, 2011). One study which used diffusion tensor imaging showed that children with ADHD have thinner, less myelinated, and less consistently organized fibers in the right cerebral peduncle, left middle cerebellar peduncle, and left cerebellum (Ashtari et al., 2005). Previous studies using MRI in children with ADHD have observed structural changes in cerebellum and specifically in the vermis (Castellanos et al., 1996; Berquin et al., 1998; Mackie et al., 2007). A section of the executive/non-motor portion of the cerebellum, the left Crus I result has been shown to have a lower grey-matter volume in ADHD children compared to healthy controls (Fernandez et al., 2022).
However, the brain changes associated with ADHD extend beyond the cerebellum. Structural brain imaging studies have revealed that patients with ADHD display smaller brain volumes in multiple regions, including the cerebellum, dorsolateral prefrontal cortex, caudate, pallidum, and corpus callosum (Seidman et al., 2005; Luders et al., 2009). Moreover, morphological and functional imaging studies in patients with ADHD have identified abnormal white-matter structure of neural pathways connecting prefrontal and parieto-occipital areas with the striatum and the cerebellum (Silk et al., 2009).
This evidence of ADHD being linked to cerebellar abnormalities is further supported by studies revealing that children with ADHD perform poorly on tasks that demand cerebellar involvement. Children with ADHD exhibit abnormal tapping responses and abnormal synchronized oscillatory mechanisms (Rivkin et al., 2003; Ben-Pazi et al., 2006). Children with focal cerebellar disorders and ADHD showed assorted impairments of eyeblink conditioning (Frings et al., 2010; Reeb-Sutherland and Fox, 2015). Ample research shows that ADHD is associated with changes in cerebellar morphology and cerebellar function (Buderath et al., 2009; Frings et al., 2010; Bledsoe et al., 2011). The role of the vermis in ADHD is still not clear but some researchers have pointed out its possible role. Bledsoe et al. (2011) highlighted that a smaller volume of the posterior inferior vermis is connected to behavioral outcomes like hyperactivity, attention and impulsivity reported by parents of children with ADHD.
Increased cerebro-cerebellar functional connectivity in the superior temporal gyrus within the somatomotor network could underlie impairments in cognitive control and somatic motor function in ADHD. Furthermore, increasing cerebro-cerebellar functional connectivity in older participants with ADHD suggests that enhanced cerebellar involvement may compensate for dysfunctions of the cerebral cortex in ADHD (Wang et al., 2022). Moreover, research also suggests that ADHD is also associated with changes in how the cerebellum interacts with other brain regions, including the cerebral cortex (Qiu et al., 2011). This may in turn explain some of the changes in cognition seen in cerebellar patients.
In summary, abnormal cerebellar development, morphological differences of the cerebellum and cerebellar connectivity, and results from behavioral studies where the cerebellum is involved prove that this structure of the brain is clearly involved with ADHD.
Neurocognitive functions in ADHDIn the following section, we analyze studies that examine the neurocognitive functions of patients with ADHD. The research indicates that patients with ADHD experience deficits across all cognitive domains, although the specific subdomains affected differ from those observed in the other disorders reviewed.
Executive functionsStudies have shown differences in neurocognitive function between patients with ADHD and controls, as evidenced by results from the 3-dimensional computerized Tower of London Test. This test measures both planning latencies and accuracy and has demonstrated that patients with ADHD exhibit impairments in both areas. This trend of impairment is thought to stem from difficulties in inhibiting responses when faced with problem-solving tasks, resulting in decreased planning activity (Young et al., 2007). The working memory capacity, which is also important for planning ability of patients with ADHD was evaluated using the Tower of London-Freiburg Version, and the results showed no significant differences between the ADHD patients and healthy controls (Mohamed et al., 2021). The Stroop Test, which measures patient’s ability to shift attention and inhibit interfering information (MacLeod, 1991), was conducted to assess the inhibitory ability of ADHD patients.
Results showed that patients with ADHD exhibit impaired inhibition compared to healthy controls, as reported in various studies (Fabio and Caprì, 2017; Guo et al., 2021; Callahan et al., 2022). However, it should also be noted that some studies have found no differences in executive control and inhibitory ability between ADHD patients and healthy controls, as indicated by similar error rates (Aycicegi-Dinn et al., 2011). While some tests suggest impairments in cognitive flexibility, attentional switching, and working memory, others find no significant differences between ADHD patients and healthy controls. For example, the Trail Making Test-Langensteinbacher Version showed no impairments in cognitive flexibility and attentional switching in ADHD patients (Mohamed et al., 2021). In contrast, adults with ADHD performed worse on Trial Making Test Part B used to measure visual attention and task switching (Tatar and Cansız, 2022). Cognitive flexibility and reaction time were measured with the Cognitive Flexibility Inventory and the Iowa Gambling Task, results were significantly lower in the ADHD patients compared to the control group (Roshani et al., 2020). Studies evaluating working memory using various tests, including the 2-Back Test, the Vienna Test System (Schellig and Schuri, 2009), and the Tower of Hanoi Test (Fabio and Caprì, 2017), also produced contradictory results. While some studies found impairments in working memory function in ADHD patients (Fabio and Caprì, 2017; Nikolas et al., 2019), others found no significant differences (Mohamed et al., 2021). The Letter/Number Sequencing Task (Gold et al., 1997) was used to assess auditory working memory, and results showed that ADHD patients had significantly lower scores compared to healthy controls (Aycicegi-Dinn et al., 2011). In contrast, visuospatial working memory, which does not require auditory processing, was found to be similar between ADHD patients and controls (Aycicegi-Dinn et al., 2011). Studies have linked deficits in executive function in ADHD patients specifically to the cerebellum. In an fMRI study, Depue et al. (2010) showed that ADHD patients with executive dysfunction had reduced cerebellar activation and decreased cerebellar volume, plus decreased activation of the frontal cortex and basal ganglia compared to healthy controls.
Another fMRI study by Hirose et al. (2014), showed changes in cerebro-cerebellar interaction during response inhibition in task performances. This suggests that the involvement of the cerebellum in the functional enhancement of inhibitory control may be sustained by modifications of the prefrontal-cerebellar interactions.
Attention and processing speedPatients with ADHD exhibit difficulties with processing speed, distractibility, and selective attention, leading to an increased number of omission errors compared to healthy controls on tests such as the Vigilance and Sustained Attention Test and Selective Attention Test, two subtests of the Vienna Test System (Sturm, 2017; Mohamed et al., 2021). Similarly, ADHD patients showed a clear impairment in the Continuous performance test which is used to measure sustained attention and response inhibition (Tatar and Cansız, 2022). Performance deficits in patients with ADHD were seen for errors of omission, correct responses, late responses, and errors of commission compared to healthy controls. Low scores emerged using the non-conflict block of the Go/No-Go task and the Stroop Color-Word Task to assess processing speed. Taken together these results have shown that ADHD patients have slower response times compared to healthy controls (Aycicegi-Dinn et al., 2011). Furthermore, research has consistently demonstrated that sustained attention, reaction time, inattention, impulsivity, response speed and variability of reaction time are significantly impaired in patients with ADHD (Tinius, 2003; Nikolas et al., 2019; Guo et al., 2021). Like with executive functions, deficits in attention and processing speed have been linked to the cerebellum. For example, structural neuroimaging studies have shown reduced cerebellar volumes in ADHD which in turn correlates with attentional problem (Petacchi et al., 2005; Kucyi et al., 2015).
LanguageThe results of verbal fluency tests in patients with ADHD are inconsistent. While some studies have found no difference between healthy controls and ADHD patients using the Resensburg Word Fluency Test (Aycicegi-Dinn et al., 2011; Mohamed et al., 2021), others have reported impaired verbal fluency when different testing methods were used (Hervey et al., 2004; Boonstra et al., 2005; Tucha et al., 2005; Guo et al., 2021). The naming ability of patients with ADHD, which is related to semantic memory retrieval capacity, has not yet been fully explored in terms of its contribution to language in this neuropsychiatric disorder.
Children with ADHD showed slower speed response than typically developed children on a naming test (Wang et al., 2017a). Furthermore, ADHD children show lower accuracy, higher number of errors and slower responses in figure naming compared to healthy controls (Koltermann et al., 2020). A previous study by Takeda et al. (2020) attempted to identify differences in Wechsler Adult Intelligence Scale profiles between ADHD patients and healthy controls, but no significant group differences were found in Verbal intelligence quotient. However, a more recent study by Doi et al. (2022) found that female ADHD patients had a lower Verbal intelligence quotient compared to males when characterizing the cognitive profile of ADHD and ASD.
The contribution of the cerebellum, together with other brain areas are necessary for language processing. Verbal fluency and lexical retrieval tasks are associated with cerebellar activation. For example, the contralateral cerebellar hemisphere, together with Broca’s area, was actively involved in the production of semantically related verbs in response to visually presented nouns. This is an example how these actions are due to non-motor cognitive processes subserving semantic word association (Petersen et al., 1988, 1989; Stoodley and Schmahmann, 2009a; De Smet et al., 2013). Further reinforcing the cerebellums role in language, Arasanz et al. (2012) demonstrated that continuous cerebellar theta burst stimulation (cTBS) using Transcranial Magnetic Stimulation (TMS) interfered with performance on phonemic and semantic fluency tasks.
MemoryADHD patients have shown lower scores in delayed recall of verbal memory compared to healthy controls as tested by the Verbal Learning and Memory Test (Mohamed et al., 2021). Likewise, immediate and delayed verbal recall, tested using the Logical Memory Subtest of the Wechsler Memory Scale, have been found to be clearly impaired in ADHD patients (Aycicegi-Dinn et al., 2011; Callahan et al., 2022). Further, the immediate and delayed visual recall tested using the Rey-Osterrieth Complex Figure test showed that ADHD patients performed worse than healthy controls (Aycicegi-Dinn et al., 2011). On the other hand, Dinn and colleagues found that patients with ADHD do not have a memory loss once the information is encoded (Aycicegi-Dinn et al., 2011).
A meta-analysis investigating the presence of procedural sequence learning found that ADHD patients do not exhibit significant deficits and that procedural sequence learning appears to be intact in ADHD (Sanjeevan et al., 2020). However, another study reported impaired results for procedural memory (Butzbach et al., 2019).
The cerebellum also contributes to memory functions, for example, Fastenrath et al. (2022) show that primarily the amygdala and successively the cerebellum is part of a network implicate in emotional enhancement of episodic memory and as well the right cerebellum is involved in semantic memory task (Gatti et al., 2021).
Visuospatial perception and visuospatial abilityVisuospatial ability, tested with the Rey-Osterrieth Complex Figure test, did not show significant differences between healthy controls and ADHD patients (Aycicegi-Dinn et al., 2011; Callahan et al., 2022). Schreiber et al. (1999) suggested that lower results on Rey-Osterrieth Complex Figure test copy trial was due to executive dysfunction in adults with ADHD and Rey-Osterrieth Complex Figure test can be a sensitive test to detect these deficits.
Social cognitionSocial cognition is still poorly explored in patients with ADHD, in particular regarding adults (Morellini et al., 2022). Contrasting results in the literature emerge for this cognitive domain. Some studies demonstrated that emotion recognition and processing, emotional prosody and empathy are impaired in ADHD patients. Control groups performed better than ADHD patients on the Cambridge Behavior Scale, the Tübingen Affect Battery and Reading mind from the eyes test (Uekermann et al., 2010; Kis et al., 2017; Tatar and Cansız, 2022). In contrast, healthy controls and ADHD patients did not differ on theory of mind task like the revised version of Reading the Mind in the Eyes Test and the Object Alternation Test (Aycicegi-Dinn et al., 2011).
Cerebellum plays a fundamental role in identifying sequences in movements that are important to understand the intentions of others. For example, mirroring, attribute mental states in others, mentalizing, and making predictions regarding social behavior (Van Overwalle et al., 2020). Interestingly, the same functions are impaired in patients with ASD diagnosis (see below).
General intelligenceIntelligence is not impaired in patients with ADHD. Researchers who used the Cattell’s Culture Fair Intelligence Test found no difference between healthy controls and adult ADHD patients (Aycicegi-Dinn et al., 2011). Likewise, Takeda et al. (2020) showed that intelligence quotient was not impaired in adults with ADHD, although working memory and processing speed indices of Wechsler Memory Scale-III were significantly lower than healthy controls.
SummaryAdults with ADHD exhibit clear impairments in multiple cognitive domains that impact their daily functioning. The cognitive heterogeneity among adults with ADHD may be due to variations in comorbid disorders, medication status, and mood disorders within the ADHD group (Cumyn et al., 2009; Chen et al., 2018).
An overview of the neurocognitive functions and how they are affected in ADHD according to this review are illustrated in Table 1. Executive functions, such as the ability to inhibit, cognitive flexibility, as well as the processing of auditory information, are particularly compromised in these patients. On the other hand, visuospatial working memory is less impacted than auditory working memory as it does not require additional auditory processing (Aycicegi-Dinn et al., 2011).
Several deficits were identified under the attention cognitive domain and processing speed as well ias verbal fluency tasks (Hervey et al., 2004; Boonstra et al., 2005; Tucha et al., 2005; Guo et al., 2021). Visual episodic memory appears to be less affected compared to verbal episodic memory in patients with ADHD. However, impairments in verbal delayed recall and visual immediate and delayed recall have been observed, but recognition ability appears to be preserved irrespective of modality (Aycicegi-Dinn et al., 2011; Callahan et al., 2017; Butzbach et al., 2019; Sanjeevan et al., 2020; Mohamed et al., 2021). Hence, the ability to store (transfer?) information to the declarative long-term memory is not affected, but rather the retrieval of information.
The results of social cognition studies in patients with ADHD are contradictory, and the presence of undiagnosed comorbid ASD could be a confounding factor that negatively impacts their social cognition abilities (Uekermann et al., 2010; Aycicegi-Dinn et al., 2011; Morellini et al., 2022).
Mental rigidity, a common feature in patients with ADHD and ASD, could also be contributing to the varying results. The presence of disinhibition, difficulties with planning, and lack of motivation, as well as other dysexecutive symptoms characteristic of frontal lobe syndrome, have been reported among patients diagnosed with ADHD. These symptoms can impact other cognitive domains, and self-report questionnaires have shown high scores for executive dysfunction, indicating deficits in neuropsychological assessments (Aycicegi-Dinn et al., 2011).
When it comes to ADHD, the presence of undiagnosed comorbidities and other psychiatric disorders can significantly impact the results of neurocognitive assessments. Additionally, the consumption of psychotropic substances and multiple medications by some ADHD patients can greatly impact their cognitive and executive function performance (Vik et al., 2004). For instance, in a study done by Crunelle et al. (2013), ADHD patients that used cocaine had higher levels of motor and cognitive impulsivity compared to healthy controls.
Collectively, the evidence from behavioral and neuroimaging studies, indicates that ADHD is associated with cerebellar abnormalities and that these cerebellar abnormalities impact cognitive functions. This suggests that the symptoms associated with ADHD are partly due to changes in cerebellar function. However, it is also clear that ADHD is a disorder that affects multiple brain regions, and more research is required to determine the precise nature of the relationship between these deficits and the extent to which they are due to cerebellar dysfunction or problems with the interplay between various brain regions, including the cerebellum. Identifying the specific contribution of cerebellar impairment in the context of ADHD is crucial to advance our understanding of this disorder and inform the development of effective interventions to improve cognitive and motor functions.
Autism spectrum disorderAutism spectrum disorder is characterized by difficulties with social interaction, communication, limited interests, and repetitive and restrictive behavior (Hodges et al., 2020). These problems can be of varying degrees and be associated with disorders of language development, general intelligence, emotional regulation, sensory sensitivity, and executive abilities. Sometimes intelligence and general cognitive functioning are preserved (Robertson and Baron-Cohen, 2017; Cai et al., 2018; Demetriou et al., 2018). There is no pharmacological treatment for ASD. However, early diagnosis and intervention coupled with remedial services can benefit patients (Baio et al., 2018).
The most common suggested causes of ASD are metabolic and physiological disorders, immunological dysfunction, oxidative stress, and mitochondrial dysfunction (Nardo
留言 (0)