1. IntroductionObesity and overweight are two of the biggest health concerns worldwide, with one-third of the population being overweight or obese [
1]. The increase in available high-fat and high-sugar food choices has contributed to increased obesity rates [
2]. The consumption of these foods and drinks has spiked in recent decades and has been associated with an increase in adiposity, biomarkers of metabolic syndrome (MetS), and associated pathologies [
2,
3].Studies have tried to elucidate how an obese phenotype impacts sweet taste. It has been reported in humans that a high BMI is associated with lower perceived sweetness, and with increased liking for sweetness [
4]. Preclinical studies have shown that the adipokine leptin has been linked to sweet taste function at several levels, from the taste receptors in the taste buds to the brain reward processing system [
5]. It is known that leptin alters sweet taste transduction through the Ob-Rb leptin receptor located in the T1R3 cells of the taste bud that are activated by sweet stimuli [
6]. In nonobese humans and rodents, the behavioral responses to a sweet stimulus change with the circadian rhythms of circulating leptin, whilst, in obese individuals, these circadian changes in leptin are absent [
7].Clinically, obesity is treated by improving diet quality and reducing caloric intake, by increasing physical exercise, and by bariatric surgery in severe cases [
8]. These interventions have been shown to affect the sweet taste function with contrasting outcomes. For example, weight loss induced through dieting [
9] or cognitive therapy [
10] has been reported to decrease liking for concentrated sucrose solutions in women, although no differences were observed in other studies [
11]. Physical exercise appears to have differential effects on sweet taste depending on whether the training is acute or chronic. Acute bouts of intense aerobic exercise increased the intake of sweet solutions and the reported liking of sucrose in healthy subjects [
12,
13], suggesting a compensatory effect of the nutrient depletion due to the exercise bout [
14], while an increase in self-reported physical activity over 6 months decreased the perceived intensity of sweet solutions [
15]. However, it is unclear whether these changes in sweet taste function are a direct result of the exercise itself or of the weight loss associated with it [
14]. Nutritional state appears not to be a strong determinant of sweet taste function [
16], and studies comparing sweet taste detection thresholds between individuals consuming vegan, vegetarian, or omnivorous diets for decades [
17] or low-sugar diets for months [
18] did not show altered thresholds. However, it has also been reported that a low-vegetable meal increases sweetness desire 2 h after meal consumption [
19], while another study reported no differences in sweet intensity and hedonics after a meal [
20].Several animal models have been developed to study obesity [
21]. Among them, the cafeteria diet (CAF)-induced obesity (DIO) model, in which animals are fed several high-fat and high-sugar items of human consumption [
22,
23], is considered a robust model with high face validity for human obesity [
24]. CAF diet induces obesity through hedonic eating and hyperphagia due to the high palatability of its ingredients; additionally, it generates more severe MetS symptoms than other high-fat and high-sugar diets [
25,
26].In animal models of DIO, a decrease in motivational behavior toward sweet stimuli, as measured by increased latency to approach the sweet stimuli and/or decreased consumption, has been reported in males [
27,
28,
29]. It has also been reported that diet-induced weight loss in male rats increases preference for low concentrations of sucrose [
28]. Furthermore, a decrease in preference for highly concentrated sweet solutions in males has been reported in models of gastric bypass surgery [
30,
31]. The effects of exercise in animal models and how it affects sweet taste have not been extensively studied. We previously reported a decrease in sugared milk and carbohydrate intake in female Sprague-Dawley rats after 8 weeks of treadmill exercise [
32]. Other authors have shown that female mice selectively bred for a high runner phenotype showed decreased consumption of artificial sweeteners when exercised [
33]. Sex differences exist in human taste perception, with women being generally more sensitive than men [
34,
35,
36]. This sexual dimorphism is also present in rats, in which females show higher intakes of sucrose solutions than males [
37,
38].The responses to sweet stimuli in animals can be interpreted in terms of wanting and liking, which are considered two separate processes [
39]. Wanting is considered to be related to how much of a reward is consumed, while liking is related to hedonic impact and to which reward is consumed [
40]. Wanting is thought to be a process mediated by the dopaminergic system that encompasses several brain structures processing reward value, such as the mesocorticolimbic dopaminergic system. On the other hand, liking is thought to be mediated by small and localized areas, termed hedonic hotspots, mainly modulated by the opioidergic system and overlapping with areas controlling wanting [
39]. These two processes are relatively independent, since it has been shown that mesolimbic dopamine depletion impairs the ability to exert motivational effort while not affecting hedonic reactivity [
41], and that elevated dopamine levels increase motivational effort while not affecting hedonic reactivity [
42]. However, while wanting can be affected without altering liking, modifications that alter liking almost always alter wanting [
40,
43].
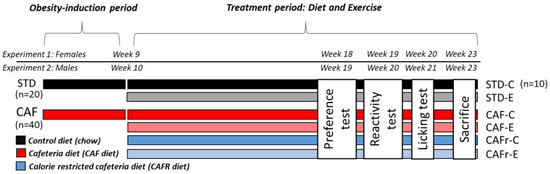
In this study, we aimed to (i) characterize the effects of CAF DIO upon the sweet taste in terms of wanting and liking, (ii) determine the putative changes in wanting and liking of sucrose solutions due to dietary and exercise interventions based on a calorie-restricted CAF diet (CAFR) and treadmill training in DIO animals, and (iii) analyze the changes in food intake, as well as biometric and metabolic parameters, induced by these interventions. We aimed to determine these parameters in Long-Evans rats, both in females (Experiment 1) and in males (Experiment 2).
4. DiscussionIn this study, we showed that CAF diet successfully induced obesity both in females and in males, and that CAFR intervention successfully ameliorated this effect. The outcomes of this dietary intervention were greater in females than in males. The CAFR diet in females decreased BW, Lee index, and adiposity and improved metabolic risk factors (decreased serum glucose, insulin, triacylglycerides, and insulin resistance), while CAFR in males decreased only adiposity and leptin levels [
44]. CAFR in both sexes decreased the proportion of non-chow foods consumed and increased the proportion of chow consumed, suggesting a healthier food intake pattern. Exercise decreased cholesterol levels in males indicating an amelioration of the MetS risk, but it did not exert noticeable effects upon the biometric variables in either sex.Regarding the biometric parameters, the obesity induced by CAF diet in this study was similar to that induced in previous studies by our group and others [
22,
29,
53]. Both sexes exhibited similar changes over the obesity induction period, with differences in BW between groups appearing earlier in females than in males. This might be explained by the fact that females presented a higher CAF intake and a lower chow intake than males, which has also been reported in a model of high-fat diet (HFD) [
54]. Previous studies reported that rodent males under HFD gained body weight more rapidly than females, which presented a delayed response [
54,
55]. These results partially contrast with ours; however, the fact that these studies used a HFD while ours used the CAF diet might partially explain the different results we obtained here. Eating patterns differ between DIO diets, with CAF inducing higher intakes than HFD [
23,
26] and reducing the amount of chow consumed due to the higher preference for palatable foods. Interestingly, females showed a higher preference for cafeteria diet overall than males, in accordance with other studies [
54]. Another interesting finding of the present study was the differential effect of CAFR diet on decreasing BWG in females but not in males, despite CAFR females maintaining a higher proportion of non-chow food intake than CAFR males. It has been previously reported in mice that, when switching from HFD to chow, females lose body weight faster than males [
55], which is in accordance with our results, since our animals were switched from CAF to CAFR diet, which increased chow intake. The CAFR diet shifted the intake of the animals toward a healthier profile that included increased chow intake, which is considered a healthier food choice than CAF, and decreased cafeteria diet intake in both sexes, which is consistent with previous observations in animals fed this diet [
29,
56]. This might partially mimic the human behavior of eating small portions of palatable energy-dense food while following dietary treatments aimed at increasing the amount of healthy food consumed (here represented by chow). This shift in intake only ameliorated biometric parameters in females, but not in males, despite the adiposity being decreased in both sexes. Metabolically, our CAF intervention resulted in a profile that can be considered characteristic of obesity (hyperglycemia, hypertryacilglyceridemia, hyperinsulinemia and increased insulin resistance, and hyperleptinemia). These consequences of CAF diet have already been widely reported by us and others [
25,
26,
29] and are indicative of the development of MetS in these animals. Regarding the metabolic and hormonal parameters, the effects of CAFR diet seemed to differ between sexes, with females showing an attenuation in the levels of circulating glucose, TAGs, leptin, and insulin, as well as the HOMA-IR, while, in males, only the cholesterol levels decreased as a result of CAFR diet [
44]. This might be explained by the greater responsiveness that females present to CAFR dieting.Exercise did not modulate the intake, and no differences were observed between exercised and control animals in food intake, this is also consistent with other reports in which exercise of a similar intensity was unable to modify energy or food intake [
47,
57]. In line with this, we also detected that exercise had no noticeable effect on either the biometric parameters or the metabolic and hormonal profile in either sex. While similar interventions to the one used here have shown a 25% reduction in body weight gain in adult Sprague-Dawley rats [
57], it seems that greater intensities or voluntary exercise rather than moderate treadmill training are more effective at inducing weight loss [
58,
59]. Another factor that might influence this parameter is the choice of rat strain; in the present study, Long-Evans animals were used, while, in Leigh et al. (2020) [
57], Sprague-Dawley animals were used. The Long-Evans rat strain is known to be more active than Sprague-Dawley [
60]; this might imply that higher exercise intensities were necessary in Long-Evans than in Sprague-Dawley in order to generate significant changes. Even in the absence of effects on those variables, exercise produced new and noticeable behavioral outcomes, decreasing sucrose consumption in both sexes and decreasing the palatability of sucrose in males.In this study, we performed three tests which act concurrently to provide information about several aspects of ingestive and motivational behavior: the two-bottle sucrose preference test, the taste reactivity test, and the brief-access licking test. In the preference test, a reduction in sucrose intake can be interpreted as anhedonia and decreased appetitive behavior [
61]. Similarly, in the taste reactivity test and the brief-access test, the number of animals initiating the behavior and the number of trials initiated, respectively, can also be interpreted as a measure of appetitive behavior [
49]. In the taste reactivity test, the hedonic reactions elicited by the tasting of sucrose are considered a measure of how much the animals liked the stimulus [
40,
46], and a similar interpretation can be made from the preference index in the preference test [
40]. Lastly, in the brief-access test, the lick responses at each sucrose concentration can be interpreted as a measure of the ingestive or consummatory behavior elicited by the tastant [
47,
48,
49]. The length of the licking episodes performed at each sucrose concentration is a measure of the orosensory feedback power and the perceived palatability of that sucrose concentration [
52,
62].The behavioral results indicated that “wanting” (i.e., how much of a reward is consumed) was affected by CAF diet. This was observed as a decrease in sucrose intake in the sucrose preference test in both sexes, a decrease in trials initiated in the brief-access test, and fewer CAF-fed animals tasting the stimulus in the taste reactivity test, compared to the STD-fed animals. These three results revealed a decrease in approaching behavior to the rewarding stimulus [
49], which is consistent with previous reports in animals fed a CAF diet [
27]. This decrease in approaching behavior in the brief-access and taste reactivity tests could result in fewer approaches in the preference test, which could explain the lower sucrose consumption. Altogether, this could be interpreted as diminished appetitive behavior, in part dependent on the dopaminergic system [
63]. Concurrent with our results, HFD induced obesity in Long-Evans rats, impaired mesolimbic dopamine function, and decreased the number of operant conditioned responses to sucrose [
63]. Moreover, chronic HFD in rats was related to a downregulation of dopamine and opioid receptors in the mesolimbic system [
64,
65]. Additionally, downregulation of D1 and D2 receptors was observed after chronic CAF diet, and this effect was greater in females than in males [
66]. Dopamine release in the nucleus accumbens (NAcc) was shown to be increased after sucrose consumption in sucrose-naïve rats [
67]. HFD diet decreased tyrosine hydroxylase-positive neurons in the dopaminergic nigrostriatal pathway [
68], and a decrease in dopamine release in the NAcc has been described after intake of milk enriched with 5% fat [
69]. The decreased expression of dopamine receptors coupled with the decrease in dopamine synthesis and release in obese animals might imply a devaluation of the reward value of sucrose [
70]. Another mechanism that might reduce sucrose intake in the preference test is the development of a depressive state that induces anhedonia in obese animals [
71,
72]. In fact, in a modified version of the sucrose preference test, in which animals are administered 1% sucrose (0.03 M) for 16 h, a diminution in sucrose intake is used to measure anhedonia [
61]. In obese rats, the decrease in sucrose intake in a sucrose preference test was interpreted as anhedonia and related to increased adiposity and leptin levels [
73]. In humans, obesity and depression are highly comorbid [
74], and weight loss due to calorie-restricted diets has been reported to improve depression scores [
75]. Several mechanisms have been proposed to link obesity and depression, including a mild level of chronic inflammation characteristic of obesity, chronically increased proinflammatory interleukin (IL-2) levels, and leptin signaling [
76,
77].
The results regarding appetitive behavior in CAFR-fed animals showed increased sucrose consumption in the preference test and more trials initiated in the brief access licking test compared to CAF diet only in females. This might imply a functional improvement in the dopaminergic system and a partial reversal of wanting compared to CAF diet. In males, there were no effects of CAFR diet on appetitive behavior, with the values of these parameters being similar to those of CAF-fed animals. Therefore, females seemed to be more responsive to the dietary treatments than males regarding appetitive behavior.
This recovery of appetitive behavior in CAFR-fed females might be related to the amelioration of the obese phenotype. Of note, people in the process of weight loss through calorie-restricted diets showed an amelioration of depression scores [
75]; in another study, an improvement of depression scores was positively correlated with a decrease in body weight [
78].With regard to the exercise intervention, a new finding to our knowledge is that treadmill exercise decreased sucrose intake (i.e., appetitive behavior) in the preference test. This result is in line with several studies indicating that exercise might produce functional changes in the dopaminergic mesocorticolimbic pathways that make the animals less susceptible to the rewarding effects of drugs of abuse [
79,
80]. For example, voluntary exercise on a running wheel decreased self-administration of cocaine in rats [
79], and the treadmill exercise in mice decreased MDMA conditioned place preference [
81]. In the same way, exercise might have similar effects on sucrose intake, being protective against overconsumption of sweet drinks since the intake of both sucrose and drugs of abuse activates similar pathways in the brain [
82]. Exercise has also been reported to increase D2R expression and protein levels in the NAcc [
83,
84], although conflicting reports have been published [
85]. In older humans undergoing regular aerobic exercise at 40–80% of maximal heart rate (walking, jogging, or cycling) for 6 months, D2R availability, measured by PET scanning, was positively correlated with VO2 max [
86], indicating that physical fitness improves dopaminergic function. Additionally, treadmill exercise at a similar intensity (15 m/min, 40 min) to the one used here induced an increase in dopamine release in the NAcc after voluntary intake of milk enriched with 5% fat in HFD rats [
69]. Regarding liking (i.e., hedonic impact or which reward is consumed), we detected a decrease in sucrose liking in CAF-fed animals, as evidenced by decreased sucrose preference in the two-bottle preference test and decreased hedonic reactivity in the taste reactivity test. This effect was greater in males than in females since males presented decreased preference in more concentrations than females, and CAF females did not present a significant reduction in positive hedonic reactions. As stated above, liking is thought to be mediated mainly by the opioid system [
87] and to be controlled by small regions in the brain [
39] termed hedonic hotspots. It has been shown that chronic CAF diet induces a decrease in opioid receptors in the NAcc in males but not in females [
66]. In Fam et al. (2022) [
27], a similar CAF diet intervention resulted in no changes to liking of sucrose. However, the intervention in Fam et al. (2022) [
27] started in adult rats, while ours started post weaning, which might indicate that changes in liking are only present when CAF diet is started early in life.
Regarding the CAFR diet, results showed no effects of this intervention on liking since both preference and hedonic reactions were not different between CAFR and CAF animals.
Exercise did not affect the hedonic reactivity in the taste reactivity test. The apparent reduction seen in CAF exercised animals compared to CAF controls can be explained by the lower number of CAF exercised animals that initiated behavior, skewing the result toward lower hedonic reactivity. This lack of effectiveness of exercise upon hedonic reactivity is consistent with previous reports of treadmill exercise having a minimal impact on opioidergic signaling [
88], considered crucial in the processing of liking [
39].
Molecular determinations of the dopaminergic and opioidergic systems are beyond the scope of this work; however, future research should emphasize this aspect, especially in animals fed the CAFR diet.
Consummatory behavior can be evaluated in the brief-access test through the lick score (difference between licks at each concentration and licks to water). Results showed that the CAF diet in males decreased the lick score at intermediate concentrations (0.1 M), with no changes at higher concentrations. This might imply that obese animals present lower consummatory behavior at these concentrations than nonobese animals, putting obese animals at risk of overconsuming at higher concentrations. CAFR diet reverted this by increasing the lick score at the 0.1 M compared to CAF-fed animals, aligning their phenotype more closely with the STD-fed condition.
We detected that exercise decreased the lick score; this effect might act in conjunction with the decreased appetitive behavior in exercised animals explained above to provide additional protection against overeating in exercised subjects.
Perceived palatability of sucrose in the brief-access test is evaluated through the burst size. This parameter is considered to indicate the potency of the orosensory positive feedback and to reflect perceived palatability [
52,
62]. We found a decrease in burst size in CAF-fed animals and more specifically in the CAF females, indicating a decreased perceived palatability in obese animals. One possible mechanism that might explain this decrease in palatability is a dysregulation of leptin signaling. The leptin receptor is present in the neural pathway relaying taste information from the tongue to the SNC. It has been shown that leptin administration induces hyperpolarization of taste receptor cells [
89] and suppresses the response of the peripheral taste nerves to sucrose [
90]. Our results showed that serum leptin levels increased in CAF groups, to a greater degree in females than in males (653% increase in females and 224% increase in males compared to the corresponding STD group). This difference in leptin levels between sexes suggests that the suppression of the signal transduction for sweet taste could be greater in females than in males. This would be consistent with the greater decrease in perceived palatability we detected in the brief-access licking test in females than in males. Regarding the effects of the CAFR diet, our results show that this intervention corrected the effects of CAF diet on the perceived palatability in females, with the burst size increased in CAFR females compared to CAF females, but not in males. This result is also consistent with the different serum leptin levels in both sexes since the decrease in them was greater in CAFR-fed females than in males (57% versus 83% reduction compared to the corresponding CAF group), suggesting an attenuated suppression of the signal transduction for sweet taste in females [
89,
90].Studies in humans have reported that, in women, weight loss induced through either low-fat or low-carbohydrate diets shifted liking for sweet food toward lower concentrations compared to baseline [
9]. In another study, obese women instructed to reduce their total caloric intake and to increase their physical activity decreased BW and BMI and shifted their perceived palatability of sucrose toward lower concentrations [
10]. Considering these results in humans, although the CAF intervention was interrupted in the females’ experiment, our animals cannot be considered in a fasted state during the behavioral testing since all animals had access to food. Furthermore, although the CAF and CAFR diets varied in the amount of food provided, they had the same food items and a very similar macronutrient composition, indicating that the diet composition per se should not be a determinant of sweet preference or liking. The results from human studies might suggest that weight loss rather than changes in feeding habits are necessary to revert the effects of the obese phenotype on perceived sweetness.Furthermore, exercise also affected the perceived palatability of sucrose, increasing it. Although this effect was present in both sexes, it was only significant in males, in which exercised animals presented the highest burst size for the 0.6 M solution, while, in controls, the highest burst size was for the 1 M solution. As stated above, leptin is a suppressor of sweet taste [
89,
90], and the fact that exercise in CAF males increased serum leptin levels might explain the increased detection threshold. This result indicates that exercise may change palatability and switch it toward lower sucrose concentrations, which would be in line with the reduced sucrose intake observed in the preference test, suggesting a protective mechanism of exercise against overeating.
One of the limitations of this study was the lack of a mechanistic characterization of the processes mediating the reported effects. Future studies should aim at characterizing the effects of CAFR and the combined CAFR and exercise intervention on the dopaminergic and opioidergic systems, both at rest conditions and as a response to sucrose intake, as well as leptin receptor expression and dysregulation of leptin signaling in the neural pathway relaying sweet taste from the tongue to the SNC.

留言 (0)