1. IntroductionWith the current SARS-CoV-2 pandemic, humans have now experienced three coronavirus (CoV) outbreaks in the last 20 years (SARS-CoV-1, MERS, SARS-CoV-2), and additional coronavirus outbreaks are expected in the future. Animal model systems are powerful tools to uncover mechanisms of pathology and to identify genetic dependencies for viral disease. Compared to non-human primates, ferrets, and other less widely used rodents, laboratory mice are the gold standard animal models of virus infection and disease course, in part as they permit genetic screens aimed at identifying immune factors and pathways required for disease and recovery. Complicating animal studies though, the ancestral strain SARS-CoV-2 did not naturally infect mice at the time of emergence, which was true for MERS and SARS-CoV-1 as well [
1,
2]. For SARS-CoV-1 and SARS-CoV-2, this host restriction is due to sequence polymorphisms in murine Ace2 [
3], the main receptor that binds viral Spike protein and permits cellular entry. To address this problem, researchers have used transgenic mice expressing human ACE2 under the control of the keratin 18 promoter (K18-Ace2). These mice can be infected with ancestral SARS-CoV-2 and subsequent viral variants wherein they can develop severe pulmonary disease [
4,
5]. However, due to ectopic expression of ACE2, these mice suffer from lethal neuroinflammation, a feature that is not seen in humans with COVID-19. Moreover, the hAce2 model complicates host genetic screens as knockout strains of interest must first be bred to the K18-Ace2 line. These limitations drove work to develop SARS-CoV-2 infection models in wild type (WT) mice using mouse-adapted (MA) viral variants [
3,
6,
7,
8,
9,
10,
11,
12,
13]. While use of mouse adapted viruses allows infection of wild type mice, it must be noted that these strains harbor mutations that can alter disease pathology such that it no longer resembles the strain of interest. Thus careful studies are needed to verify the effectiveness and suitability of these strains as model organisms. Studies employing mouse-adapted SARS-CoV-2 strains have primarily been performed in BALB/c mice [
6,
8,
9,
13,
14] and focused on a single sex or single age cohorts [
3,
10,
12,
15,
16], or have employed highly adapted and virulent variants that limit the ability to comprehensively examine infection and recovery [
11].The SARS-CoV-2 pandemic is moving towards an endemic phase and is likely to remain a persistent threat as the viral genome continues to evolve [
17]. Viral evolution of SARS-CoV-2 leads to the continual emergence of new variants of concern replacing previous circulating variants [
18], and also led to the ability of SARS-CoV-2 variants to infect mice [
19,
20,
21]. Additionally, current SARS-CoV-2 variants mediate a disease course that is much less pathogenic than the ancestral strain and has transitioned from causing a severe lower respiratory tract infection to a relatively mild upper respiratory tract infection [
17]. Despite the lessened pathogenicity of newer SARS-CoV-2 variants, there remains a need to understand the processes underlying infection and recovery. Moreover, it is important to note that despite the potential for deadly outcome of severe COVID-19, ~99% of people infected with SARS-CoV-2 throughout the pandemic recovered from infection with only mild to moderate disease or were asymptomatic (
https://covid19.who.int/, accessed on 1 October 2022). Thus, there remains an important need for comprehensive in vivo analyses of SARS-CoV-2 infection models to understand the dynamics of infection and recovery. For example, there is a clear and pronounced increase in disease severity based on age and sex [
22,
23,
24], but it is not fully understood whether these differences are driven by age- and sex-dependent comorbidities (such as obesity) or inherent differences of the host (such as sex-linked expression of immune genes and sex-hormone sensitive immune cells). In vivo models of infection can set the stage for future genetic screens aimed at identifying key immune pathways required for recovery as well as identifying the age- and sex-based factors that affect disease severity. Therefore, the goal of the present study was to develop an immunocompetent mouse model of SARS-CoV-2 infection that displays main features of human infection and recovery, starting with ancestral SARS-CoV-2. Here, we comprehensively describe a C57BL/6 model of SARS-CoV-2 infection and show the utility of this model in examining acute infection, progression to disease, lung disease, and recovery. As a great many transgenic C57BL/6 lines exist that lack specific genes or express modified versions of specific genes, we demonstrate that this model provides a platform for genetic interrogation of the virus/host interactions that control SARS-CoV-2 infection and immunity. 2. Materials and Methods 2.1. Mice10 wk-old C57BL/6J mice (Jax #000664) were purchased from Jackson laboratories directly or bred in-house from animals purchased from Jackson laboratories. 20-wk old C57BL/6J mice were purchased from Jackson Laboratories at 10 wks of age and aged to 20 wks old. SCID (B6.Cg-Prkdcscid/SzJ, #001913) mice were purchased from Jackson laboratories. 2 yr-old C57BL/6 mice were obtained from the National Institute of Aging subsidized aging rodent colony at Charles River, Inc. Mice were housed with appropriate food, water, and enrichment in individually ventilated cages and were euthanized if they fell below 70% original weight or became otherwise moribund, consistent with previously published studies [
3,
8]. Weights of all animals studied and numbers of mice per cohort in each study can be found in
Supplementary Table S1; however, studies of weight loss used 5 mice per cohort and tissue collection studies used 3 mice per cohort unless otherwise noted. 2.2. Infections and Monitoring
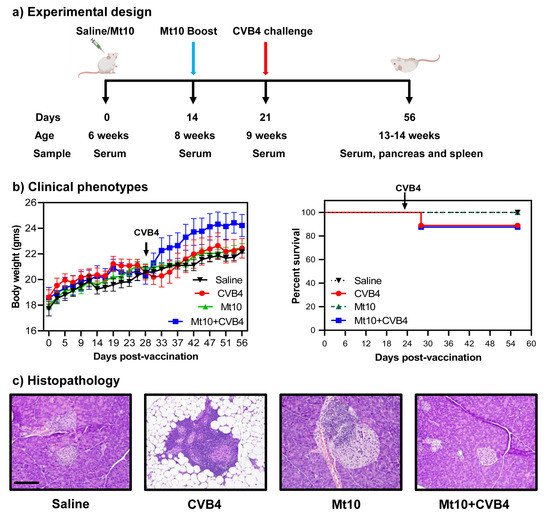
Mice were anesthetized with Ketamine/Xylazine at roughly 80–100 mg/kg ketamine and 5–10 mg/kg xylazine. Anesthetized mice were inoculated intranasally with indicated doses of virus diluted in saline at a total volume of 50 mL or with 50 mL saline alone as a mock control. Mice were closely monitored following anesthetization until they recovered and then were monitored daily for changes in health and/or weight. Mice were also monitored daily by UW DCM staff to ensure the animals always had sufficient food and water and that cage conditions met approved standards.
2.3. Virus
SARS-CoV-2 MA10 was kindly provided by Ralph S. Baric. A P3 stock was amplified to 8 × 106 PFU/mL in VERO (USAMRIID) cells and sequence verified. Sequence analysis indicated an R2G mutation in NSP7.
2.4. RNA Prep
Tissue collected at necropsy was originally placed in RNA later (Thermofischer, Waltham, MA, USA, AM7021), stored at 4° for 24 h to one week, and then transferred to −20° for longer storage. To process tissue, samples were transferred to 1 mL Trizol (Thermofischer, 15596018) in 2 mL Percellys hard tissue homogenization tubes (Cayman Chemical Co., Ann Arbor, MI, USA, 10011151). Samples were homogenized in a Percellys 24 Homogenizer using 1–3 cycles of 15” at 6500 rpm and placing samples on ice between cycles. RNA was then purified from the Trizol homogenate using the RNeasy 96 kit (Qiagen, Hilden, Germany), according to manufacturer’s protocols.
2.5. Histology & RNA-ISHFor each mouse a sample of lung was incubated in formalin at room temperature for at least seven days to fix tissue and inactivate virus. The fixed tissue was processed and embedded in paraffin. 5 µm sections were cut and stained with hematoxylin and eosin (H&E). Airway pathology was assessed in H&E stained sections to assess bronchial epithelial cell death (score: 0 = no dead cells, 1 = 1–5 dead cells, 2 = 6–10 dead cells, 3 = 11–20 dead cells and 4 = >20 dead cells; scored for ten 400× fields per mouse lung); endarteritis, venulitis, and perivenule, peribronchial and periarterial inflammation (score 0 = none, 1 = 0–25% circumference with >1 leukocyte cell layer, 2 = 26–50% circumference with >1 leukocyte cell layer, and 3 = 50–100% circumference with >1 leukocyte cell layer; scored for 10 400X fields per mouse lung); and interstitial pneumonitis and pulmonary edema (score = percentage of pulmonary alveolar parenchyma with septae expanded by leukocyte; scored for ten 100X fields). Scoring sheets showing scores for individual mice as well as calculations of average scores are found in
Supplementary Tables S4–S7. Importantly, the pathologist was also blinded to samples for analysis.RNA in situ hybridization (RNA-ISH) was performed on FFPE sections of mouse tissues using the V-nCoV2019-S probe (Catalog# 848561) for sense strand of the Spike gene and the V-nCoV2019-S-sense probe (Catalog# 845701O) for the antisense strand of the Spike gene. RNA-ISH signal was developed using the RNAscope™ 2.5 HD Assay—RED development kit (Catalog # 322350) from ACDBio Inc. (Newark, CA, USA), and slides were counterstained with hematoxylin for tissue visualization. For scoring of infected bronchioles, individual bronchiole profiles were assessed for the percentage of infected cells according to the scheme outlined in
Supplemental Figure S8. Each of these bronchioles was also assessed for a shedding phenotype defined as ≥10 ISH-positive detached cells within the bronchiole lumen. 2.6. nCounter and Bioinformatic AnalysesnCounter was run according to manufacturer’s protocol and custom probe set (
Supplemental Table S8). RCC output files were assessed for technical QC flags using the nSolver software (nSolver Analysis Software 4.0.70, NanoString Technologies Inc, Seattle, WA, USA) provided by NanoString. Raw reads were next collated from RCC output files into a single count matrix in Rstudio. Because our probe set was highly non-random by design, the number of observed counts for each gene probe were several orders of magnitude greater between mock animals and those acutely infected with SARS-CoV-2. As a result of these large differences in library size, we developed a custom approach for analysis of these data that avoided artifacts introduced by distribution-based normalization methods; our methods are conceptually related to the family of log-ratio transformations that address the compositional nature of sequence-based data.
To perform these analyses, we leveraged count data within each sample vector as an internal reference, against which we could compare all genes of interest. To do so, we first measured the coefficient of variation (CV) for all genes in the probe set, which included: genes of interest (GOI), housekeeping (HK) genes, positive controls (PCs) and negative controls (NCs). Reassuringly, positive control probes were the least variable of all genes assessed, which was expected given their role in measuring technical variation of the sample prep and assay itself, both of which are demonstrated to be low. We then used the calculated CVs to select the 3 housekeeping genes with the lowest variability. To obtain our within-sample reference value, we calculated the geometric mean of the 3 selected HK genes. We then “tethered” all count data from each sample to its within-sample reference, which we refer to as tethered gene counts (TGCs).
To visualize data in PCA space and calculate vectors for biplot analyses, we utilized the log2 value of the TGCs. To assess changes in individual genes across groups, we simply used the TGC values. In order to calculate log2 fold-change, we formed a ratio using the TGC for each GOI against its age-matched values from mock animals, and subsequently took the log2 of these values (
Figure 1).
To calculate statistical significance, we first checked for normalcy of TGC within each gene/population to be tested using a Shapiro Test. Following BH correction for multiple tests, we observed all data to be normal. We subsequently performed 2-way ANOVA using age and days-post infection as variables of interest. These data were then assessed by Tukey’s post hoc correction, and thus, all reported p-values are Tukey adjusted.
4. DiscussionWe provide a model of SARS-CoV-2 infection and recovery in C57BL/6 mice, featuring the mouse-adapted R2G virus strain which is a variant of the MA10 strain. We present a robust and comprehensive comparison of early to late infection in young, mature, and elderly adult mice, both in males and females. Our analyses include assessments of virus density and localization, animal weight loss kinetics and mortality, disease pathology across multiple tissues, and innate immune responses. We show that as in humans [
29,
30,
31], disease is mild and quickly resolved in young, healthy individuals; however, disease severity is age- and sex-dependent, with older animals (especially older males) showing greater sensitivity to lower virus challenge doses, increased mortality, increased virus persistence with systemic virus spread, and greater lung damage. We show that the magnitude of the innate immune activation displayed by innate immune gene induction is greatest in young mice and that virus and the innate immune/inflammatory responses persist longer in aged mice, suggesting that these altered responses compared to young mice contribute to viral persistence and increased lung injury in elderly mice. Finally, we show through comparison of wild type and SCID mice that the innate immune response is the primary driver of weight recovery and early stage viral control in vivo, while an intact adaptive response is still required for efficient virus clearance at later infection time points. Together, these data characterize the effects of SARS-CoV-2-R2G infection in C57BL/6 mice, which are remarkably similar to those in humans [
25,
29,
30,
31], as well as what was seen for the 2003 strain of SARS1 [
35,
36]. Additionally, this study paves the way for future studies aimed at identifying key immune pathways that are required for recovery and disease pathology following infection with not only the original strain of SARS-CoV-2 but future emergent coronaviruses as well.A number of excellent groups have previously reported model systems for the in vivo study of SARS-CoV-2 infection in mice. Early in the pandemic, these studies required the use of mice expressing human Ace2 since original isolates of SARS-CoV-2 did not infect mice due to differences in human and murine Ace2 [
37,
38]. While informative, infections in these mice did not replicate human disease due to the inappropriate localization and expression of hAce2. As the pandemic progressed, several groups reported on mouse-adapted strains of SARS-CoV-2 that acutely infected mice and produced disease reflecting what was seen in humans at that time [
3,
6,
7,
8,
9,
10,
11,
12,
13,
14]. Though several of these studies examined the effects of infection in C57BL6 mice [
3,
7,
10,
11,
12] these were limited to single sex or age cohorts or employed strains of SARS-CoV-2 that were too virulent for elucidating immune mechanisms of recovery by genetic screens. Compared to the few studies in C57BL/6 mice, the most comprehensive early studies with mouse-adapted strains characterized infection in Balb/c mice [
3,
7,
11,
13] which do not permit the vast number of genetic screens that are possible with transgenic and gene knockout lines on the C57BL/6 background. Important to our future goals of genetically dissecting key immune pathways for disease recovery of knockout mouse strains on the C57BL/6 background, the immune responses and disease severity observed in Balb/c and C57BL/6 mice are different. For example, Balb/c mice are skewed towards a Type II immune response, which may exacerbate disease as in severe COVID, while C57BL/6 mice are skewed toward a Type I antiviral/inflammatory immune response, which is needed for efficient clearance of virus [
39]. This notion is supported by studies showing that Balb/c mice experience more severe disease compared to C57BL/6 mice of the same age [
3]. This and other differences necessitated the more comprehensive study of SARS-CoV-2 infection in C57BL/6 mice presented here.Interestingly, in addition to their usefulness in genetic screens, C57BL/6 mice may better reflect disease recovery in humans than Balb/c mice. Specifically, virus tropism in C57BL/6 lungs appears to more closely phenocopy that in humans, where intense staining was observed in bronchial epithelium in addition to the alveolar space. In contrast, SARS-CoV-2 does not apparently localize to the bronchial epithelium in Balb/c mice ([
3,
25] and present study). Moreover, staining of bronchial epithelium in humans and C57BL/6 mice correlates with shedding of dying cells into the bronchial lumen ([
3,
25] and present study). Whether this outcome is important in virus clearance or virus spread or in the increased disease pathology in Balb/c mice relative to C57BL/6 mice is unclear. While not necessarily exclusive to C57BL/6 mice, the prominent pulmonary edema and interstitial pneumonitis seen in the older mice in our study also corresponds with the pulmonary edema seen radiographically in humans, and suggests that this phase of injury may resemble the early stage of SARS-CoV-2 induced acute respiratory distress in humans underlying COVID-19 [
40,
41]. Our finding that infection-mediated lung injury follows clearing of the virus, and that more extensive lung injury is observed in older mice that have delayed viral clearance, suggests that a robust innate immune response in young mice leads to rapid clearing of the virus and limits the immunopathology in the lung, processes that ascertain SARS-CoV-2 infection outcome in most humans. Together, these observations suggest that C57BL/6 mice accurately capture infection and outcome dynamics observed in humans.Novel variants of SARS-CoV-2 have arisen faster than detailed studies aiming to characterize their properties can be completed before being replaced by a new emergent variant. In fact, more recent variants, including the current omicron variants, developed the ability to infect mice and rats [
19,
20,
21], which share Ace2 sequence similarity in the residues to which SARS-CoV-2 binds. What drove this adaptation is uncertain, but it is interesting that cryptic SARS-CoV-2 variants were found in the sewers of New York City that contain mutations that are found in mouse adapted MA10 and R2G as well as in omicron [
42]. However, while omicron is able to infect mice, it is important to note that omicron-induced disease is much different from that driven by the ancestral SARS-COV-2 and appears to preferentially infect upper airways as opposed to lower airways and with milder symptoms, both in humans and mice [
19,
20,
21,
43]. Moreover, while the ability of omicron to infect mice provides the very important opportunity to study natural infections of current strains in vivo, this variant does not allow studies of recovery from the more severe infections and disease pathologies engaging the lower airway that better reflect those arising from the ancestral virus and earlier pandemic variants. As with many viral infections in humans, older individuals and older males in particular are more sensitive to infection with SARS-CoV-2 and are more prone to experience and succumb to severe disease [
22,
23,
24,
44]. This is likely due to a combination of factors that include comorbidities such as heart, lung, renal, and liver disease, obesity, dementia, and cancer which are more prevalent in older individuals [
45] as well as X-lined expression of many immune genes and the contrary responsiveness of immune cells to estrogen/progesterone and testosterone [
22]. We show here, though a comprehensive analysis of the effects of both age and sex on SARS-CoV-2 infection in mice, that older mice are much more sensitive and susceptible to infection than younger mice (10-wk), consistent with what is seen in humans. Surprisingly, though elderly (2 yr) female mice are less susceptible to infection than mature adult (20 wk) female mice. The cause of this difference is uncertain but held true over multiple independent experiments within this study; however, a similar outcome was also reported for influenza A virus infection in mice [
32]. In humans, increased levels of inflammatory cytokines such as IL-6 were found in older males with severe COVID whereas increased levels of anti-inflammatory cytokines like IL-10 were observed in older females 14 days post infection than in males [
23]. Likewise, we observed increased levels of IL-6 in elderly mice by NanoString analysis innate immune gene expression, but our study did not have the power to segregate males from females, necessitating future studies to elucidate the molecular and immunologic underpinnings driving the sex-based differences to SARS-CoV-2. Importantly, these observations further validate this model system as a relevant model for the study of SARS-CoV-2-mediated disease and recovery.
In conclusion, we present a comprehensive analysis of SARS-CoV-2 infection and recovery in C57BL/6 mice, showing responses, pathologies, tropism, distribution, and age- and sex-dependent differences in disease severity that mimic what was seen following infection with the original isolates of SARS-CoV-2, when the virus first emerged in humans. These observations serve as a basis from which future C57BL/6-based genetic screens can be compared. Additionally, due to the increased reproducibility of R2G over the parental MA10, infection of C57BL/6 mice with the R2G variant of SARS-CoV-2-MA10 is an ideal model for SARS-CoV-2 mediated disease and recovery.

Comments (0)