1. IntroductionRabbit meat has recently acquired increasing popularity as a “functional food” due to its excellent nutritional and dietetic properties. It is distinguished by its content of essential amino acids and easily digestible proteins [
1]. It has lower cholesterol and sodium levels than other meats [
2], does not contain uric acid and has a low content of purines [
3]. It is also an important source of B vitamins. Consumption of 100 g of rabbit meat contributes to 8% of daily vitamin B2, 12% of vitamin B5, 21% of vitamin B6, and 77% of vitamin B3 necessities, and fulfills the daily vitamin B12 requirement [
1]. Moreover, it contains antioxidant vitamins such as vitamin E (α-tocopherol). It has a low-fat content, and its fatty acid composition may be strongly modified by changes to rabbit diet [
3,
4]. Thus, it is of high nutritional value and can even be regarded as a functional food through the manipulation of rabbit diet and supplementation with bioactive compounds [
2,
5].During the intensive production of rabbit meat, stressed animals show altered gut microbiota and immune responses, pathogenic proliferation, decreased feed conversion efficiency and a zootechnical response [
6], mainly due to digestive disorders associated with their diet. Dietary fiber is widely considered to be the most important nutritional factor for the prevention of digestive pathologies. Therefore, a balance between less digestible (lignocellulose) and digestible (hemicelluloses and pectin) fibers is recommended, as is the addition of agro-industrial byproducts (brans and pulps) with high digestible fiber content. Fibrous raw materials are the main constituents of commercial rabbit feeds. Both the physical and chemical characteristics of dietary fibers have implications for feed intake, gut health, feed efficiency and performance. The importance of dietary fibers in animal feeding is due to their influence on the rate of passage and mucosal functionality and their role as substrates for gut microbiota. Nevertheless, their impact on digestive health is still questioned; the effects of other cell wall components such as indigestible lipids (cutins and waxes) that may interfere with fiber digestibility have not been thoroughly investigated [
7]. Moreover, there is a need to study animal nutrition using a technological and low-cost approach. Research should involve nutritional evaluation of digestibility, feeding value and biohazards, and an assessment of the feasibility of waste management and utilization. Different strategies and agro-product bioprocessing methods must still be evaluated to produce animal feed in a cost-effective way, leading to sustainable eco-management [
8]. A wide variety of supplements such as plant byproducts [
9,
10], microalgae [
11], microbial biomass [
12] and enzymes [
13] have demonstrated their positive effect on productive parameters in rabbit meat production. A less explored alternative is the employment of white-rot fungal biotransformed substrates. White-rot fungi (WRF) have been used in the degradation of crop residues under solid-state fermentation (SSF), as they are reported to decompose lignin more efficiently than any other group of microorganisms [
14]. White-rot basidiomycetes produce enzymes that can break down the main components of plant cell walls, hence increasing nutrient bioavailability [
14,
15,
16,
17]. Nevertheless, to employ lignocellulose waste in animal nutrition, optimization of the fungal pretreatment is required to shorten the process of delignification and make it more selective for lignin [
18]. Experimental evidence suggests that the competition for space and nutrients may result in enhanced degradation of lignin during solid-state co-cultivation [
19,
20]. Trametes maxima CU1 and Pycnoporus sanguineus CS2 are two basidiomycetes from northeastern Mexico with great lignocellulolytic potential [
21], recognized for their production of high-redox potential laccases, as well as thermostable cellulases, xylanases and pectinases [
22]. Enzymatic extracts of T. maxima CU1 improved the productive parameters and carcass yield of rabbits [
13]. Furthermore, sorghum hydrolyzed using a co-culture of both fungi contributed to a better rabbit meat texture [
23]. The purpose of this study was to compare a rabbit diet based on untreated sorghum with a second one that included the cereal biotransformed by the co-cultivation of these basidiomycetes. Their effects on cereal, diets, and fecal structural fiber composition, as well as on rabbit intestinal morphology, were assessed. 2. Materials and MethodsThe study was performed in General Marín, Nuevo León, Mexico. Geographically, it is located at 25°52′13.5″ latitude north and 100°02′22.56″ longitude west, with an altitude of 355 MASL. Rabbit management and care protocols were performed according to the national policies laid down by the Mexican Animal Welfare Standard NOM-062-ZOO (1999) [
24].
Fungal strain
T. maxima CU1 and P. sanguineus CS2 were obtained from the culture collection of the Laboratory of Enzymology, Biology Department from the UANL. Both strains were conserved in YMGA (glucose 4 g L−1, malt extract 10 g L−1, yeast extract 4 g L−1 and agar 15 g L−1) slants at 4 °C with periodic subcultures every three months.
Sorghum biotransformation
The strains used for sorghum biotransformation were cultivated at 30 °C for 7 d in individual flasks containing potato dextrose broth (Becton, Dickinson and Company, Frankling Lakes, NJ, USA). Prior to culturing, sweet sorghum (Sorghum bicolor) grains were washed with tap water and placed in polyethylene containers for sterilization at 121 °C for 1 h. Ten milliliters of each culture medium were used to inoculate 500 g of washed and sterilized sorghum contained in polyethylene bags. The inoculated sorghum was mixed well and incubated at 30 °C for 30 d. Finally, the hydrolyzed sorghum was milled and included in the preparation of the T2 experimental diet. Grain samples were taken to determine the initial and final fiber compositions.
Experimental design
A completely randomized design was used in which 24 unsexed New Zealand rabbits (Oryctolagus cuniculus) (532 ± 65 g), weaned at 21 d of age, were distributed between two treatments: T1 (Control diet and non-transformed sorghum) and T2 (Control diet and sorghum transformed by dual SSF with T. maxima CU1 and P. sanguineus CS2). The rabbits were individually bred in metallic net cages (45 × 60 × 50 cm) fitted with a drinker and a feeder. The diets were formulated according to the NRC (1994) [
25] (
Table 1); however, in T2 sorghum was transformed by SSF with T. maxima CU1 and P. sanguineus CS2 before its use as an ingredient.
Sorghum and fecal fiber composition
The chemical composition of fiber fractions in the sorghum samples and rabbit feces was determined according to standard methods [
25,
26]. The percentage of total dry matter (DM) was determined after oven drying (VWR HAFO® 1600 series, Atlanta, Georgia, USA) at 105 °C for 24 h by the following equation:
%DM=Winitial−WfinalWinitial×100
where Winitial is the initial weight and Wfinal is the final weight resulting from oven drying.Neutral detergent fiber (NDF DM) and acid detergent fiber (ADF DM) were measured using an ANKOM2000 fiber analyzer; for this purpose, 0.5 g samples were placed in filter bags and boiled at 100 °C for 1 h in a solution of neutral detergent (sodium lauryl sulfate, pH 7.0) supplemented with Na2SO3 and α-amylase. Afterwards, the samples were rinsed 4 times with bidistilled water (70 °C) and the NDF content was calculated according to the following equation:
% NDF DM=[W3−(W1×C1)]×100W2
where W1 is the weight of the empty bag (g); W2 is the weight of the sample (g); W3 is the weight of the dried simple after boiling it with NDF solution (g); and C1 is the blank correction factor.The ADF DM content (lignin and cellulose) was determined with cetyltrimethylammonium bromide to solubilize the hemicellulose in NDF. Concentrated sulfuric acid was used to digest the fibrous portion of the ADF residue, solubilize cellulose and estimate lignin content. Hemicellulose and cellulose contents were calculated as the difference between NDF DM–ADF DM and ADF DM–ADL DM, respectively. The nitrogen (N) content of the agricultural waste was determined by the standard Kjeldhal method [
25] and the amount of crude protein was calculated as N × 6.25.The apparent nutrient digestibility coefficients of the diets were determined according to the formula depicted by Safwat [
27]:
where CAD is the coefficient of apparent nutrient digestibility, NI represents the nutrient intake (kg DM), and NE expresses the nutrients excreted (kg of feces DM).
Rejected feed and feces were collected daily and stored at −21 °C.
Effects on intestinal morphology
Intestinal morphology was analyzed following the Attia et al. protocol [
28]. Sections of approx. 1 cm were collected from duodenum, jejunum, ileum, and caecum, washed with physiological solution, and preserved in 10% neutral buffered formalin. Paraffin-embedded samples were cut and stained with hematoxylin–eosin. Intestinal villus length and crypt depth were measured. All the assays were performed in triplicate. Villus length was measured from the apical end up to the crypt–villus junction. The crypt depth was determined from the base of the crypt to the level of the crypt opening. The morphological index was determined by image processing (Version 1, Leica Imaging System Ltd., Cambridge, UK).
Statistical Analysis
Data were subjected to ANOVA using the SPSS Statistics 10.0 (SPSS, Chicago, IL, USA) software program. Analytical determinations data are the mean of three replicates. Significant differences between treatments were compared by Tukey’s test at a 5% level of probability.
3. Results and Discussion
Composition of the Fiber Sorghum and Diets
Table 2 depicts the composition of the fiber fraction of untreated sorghum (control), sorghum transformed by dual SSF with T. maxima CU1 and P. sanguineus CS2, and of the diets based on both substrates. The contents of NDF DM, ADF DM, lignin, hemicellulose, cellulose, and ash showed a decrease of 11, 41, 80, 5 and 78%, respectively, in the treated sorghum. A similar behavior was also observed in the diet supplemented with this sorghum. The extracellular enzyme systems responsible for degrading polysaccharides and lignin in plant cell walls by the action of hydrolases and oxidoreductases [
14]—produced by the white-rot fungi T. maxima CU1 and P. sanguineus CS2 [
21]—may explain these results. In a previous work, these basidiomycetes showed their potential to biotransform buffel grass (Cenchrus ciliaris) by the secretion of cellulases, xylanases and laccases, increasing nutrient bioavailability and thus nutritional quality [
16]. T. maxima CU1 is distinguished by its ligninolytic potential, while P. sanguineus CS2 mainly degraded cellulose and hemicellulose [
16]. Therefore, while co-culturing both strains, a positive synergistic effect between their lignocellulolytic enzymes may enhance the degradation of the sorghum fibrous fraction. Other ligninolytic basidiomycetes such as Pleurotus ostreatus exhibited higher cellulose- and hemicellulose-degrading capabilities when cultured in agro-industrial waste [
15,
29]. Besides the effects on plant cell wall components, an increment in the protein content of the fermented substrates, probably due to the addition of fungal protein, was also observed, showing the potential of SSF by basidiomycetes to enhance the nutritional value of lignocellulosic substrates [
29,
30,
31]. Although Oduguwa et al. reported a rise in the crude protein content of fermented corn cob, rice bran and cowpea husk, in this work crude protein content did not increase in fermented sorghum grains [
32]. These differences might be explained by the diversity in lignocellulosic substrates employed and by dissimilar fungal degradation abilities.
Fecal Fiber Composition
Rabbit body weight at slaughter did not show statistically significant differences (p > 0.05) between treatments (the average body weight was 1634.4 g) and the mortality rate was 0% for both treatments. Likewise, Ribeiro et al. evaluated dietary incorporation of olive leaves pretreated with Ganoderma resinaceum and untreated leaves and did not find differences in rabbit growth performance [
33]. In contrast, Andrade et al. assessed the potential of several white-rot fungi to upgrade the nutritive value of cowpea (Vigna unguiculata) stover for rabbit feeding [
34], finding that treatment with Pleurotus citrinopileatus led to an efficient delignification process, which resulted in higher in vitro organic matter digestibility, showing the potential of this fungus to enhance the nutritional value of this feedstock. However, the combined effect of fungi on delignification and digestibility is mostly dependent on the fungal strain, the substrate, and the extent of fungal biodegradation [
30].
Table 3 shows the fiber composition of the rabbit feces. MS content did not differ between treatments (p > 0.05). Ash content at 7 and 21 d was higher in rabbits fed with untreated sorghum (T1), whereas at 35 and 49 d higher contents were observed in rabbits fed with fungus-treated sorghum (T2). Less organic matter and lignin (p ≤ 0.05) were detected in the excrement of T2-nourished rabbits at 35 and 49 d; likewise, diminished NDF and ADF contents were detected after 21 d. Although hemicellulose percentages did not show statistically significant differences between the two tested groups (p > 0.05), its content was lower in T2 feces for all the evaluated periods. On the other hand, in T2 excrements cellulose content was inferior only after 7 and 21 d (p ≤ 0.05). These results might explain the soft consistency of T2 feces (data not shown), and they also suggest that T2 afforded better digestibility, probably in conjunction with the decrease in fiber content [
34]. Ribeiro et al. proved that pretreatment of olive leaves with G. resinaceum only improved crude protein and fat digestibility without affecting the fiber fraction [
33]. However, in this work, sorghum fiber content was modified by dual SSF with T. maxima CU1 and P. sanguineus CS2. The synergic effect between their lignocellulolytic enzyme systems might have contributed to the increase in digestibility. SSF by basidiomycetes improved the nutritional profile of cereal grains by increasing water-soluble protein and amino acid nitrogen contents, and by promoting the hydrolysis of starch into dextrins, hence enabling their assimilation in the rabbit digestive tract [
23]. Moreover, greater antioxidant activity was reported as well [
35]. Thus, cereals can be processed into functional foods containing bioactive substances after SSF by basidiomycetes.
Effects on Intestinal Morphology
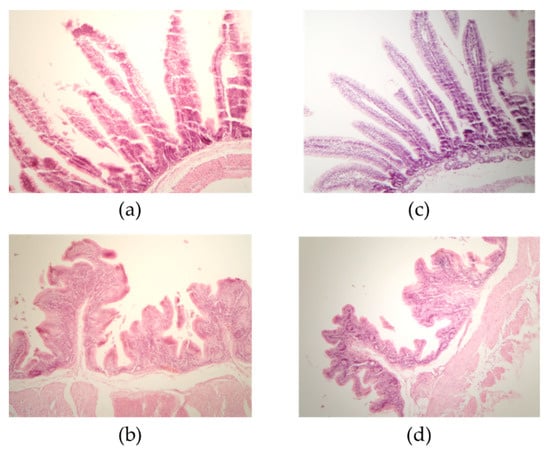
Effects on small intestine and caecum villus length and crypt depth in rabbits fed diets containing untreated sorghum (T1) and sorghum altered by SSF with T. maxima CU1 and P. sanguineous CS2 (T2) are summarized in
Table 4. Rabbits nourished with the T1 diet showed longer duodenal and ileal villi (p ≤ 0.05;
Figure 1), while no statistical differences were found between the two tested groups at the jejunum level (p > 0.05). El-Badawi et al. reported that rabbits fed Moringa oleífera-supplemented diets showed clear increases in jejunum villus height, in the villus-to-crypt ratio and in the depth of the cecal tunica mucosa, which might lead to better absorption of nutrients and microbial metabolites and to a positive effect on the feed conversion ratio and average daily gain of rabbits [
36]. Thus, the differences in intestinal morphology observed in this work might be explained by the enhanced nutrient availability in the T2 diet, which is associated with the increased accessibility to plant cell wall components due to the action of lignocellulolytic enzymes produced during dual SSF with T. maxima CU1 and P. sanguineus CS2. Likewise, Attia et al. found that supplementation with exogenous digestive enzymes in growing rabbits was associated with improved digestive enzyme activities and intestinal mucosal morphology, coupled with higher growth rates [
28].These enzymes might contribute to the hydrolysis of non-starch polysaccharides, reduce the viscosity of gut contents, and improve NDF and ADF digestibility and nutrient absorption. The morphological changes recorded in the intestinal mucosa (increased villus length and crypt depth) might be complementary changes that occurred to meet the increased rates of digestion and absorption mediated by the coupled activities of exogenous and endogenous digestive enzymes. At the cecum level, even greater differences were detected between both diets (p ≤ 0.05). The effect of SSF sorghum on cecal villi might be a response to increased nutrient availability due to sorghum pre-hydrolysis. As the digestion retention time in the caecum is relatively short, it is mainly the most rapidly fermentable cell wall polysaccharides (such as the hemicellulose fraction) that are digested at this point, playing a key role not only in rabbit digestive processes but also in the digestive health and sanitary status of rabbits [
7,
37]. T1-supplied rabbits exhibited deeper crypts in all gut tracts except for the colon (p ≤ 0.05), probably due to the increased demand in nutrient absorption, which is mainly attributable to the dietary fibers that are bioavailable after the pretreatment with white-rot fungi and their lignocellulolytic enzyme complexes [
36,
38]. Thus, supplementing growing rabbit diets with sorghum altered by dual SSF with T. maxima CU1 and P. sanguineus CS2 might contribute to increase intestinal transit time, improve mucosal functionality, and modulate their microbiota. Further investigations will be performed to explore these issues.






留言 (0)