
Remember me



Hematopoiesis and cardiogenesis are among the earliest developmental processes in vertebrates, forming the foundation of the circulatory system. In mammals, embryonic hematopoiesis occurs in multiple waves across extra- and intra-embryonic sites [1]. The first wave, known as primitive hematopoiesis, arises in the yolk sac blood islands, producing erythromyeloid progenitors (EMPs) that generate primitive erythrocytes and macrophages [2, 3]. A subsequent wave originates from hemogenic endothelial cells in the yolk sac, generating late-EMPs that migrate to the fetal liver, where they expand and differentiate into multiple hematopoietic lineages, including monocytes [4]. Concurrently, hematopoietic progenitors also emerge from intraembryonic hemogenic endothelium in the aorta-gonad-mesonephros (AGM) region, giving rise to definitive hematopoietic stem cells (HSCs) [5].
In addition to these well-characterized hematopoietic sites, recent studies have revealed that endocardial cells in the developing heart possess transient hematopoietic potential, establishing the heart as a previously unrecognized fetal hematopoietic niche. While hematopoiesis within the heart was first described in Drosophila larvae [6,7,8], where the cardiac tube functions as the primary site of blood cell production, its existence in vertebrates has only recently gained attention [9,10,11,12,13,14]. The evolutionary conservation of this process suggests that endocardial hematopoiesis represents an ancestral mechanism retained across species.
Despite its evolutionary significance, endocardial hematopoiesis has long been overlooked in mammals, likely due to its minor contribution to total blood production. However, recent findings indicate that endocardial-derived hematopoietic progenitors give rise to macrophages that play a unique and indispensable role in cardiac morphogenesis—functions that cannot be compensated for by macrophages originating from other hematopoietic sources [10, 13]. These discoveries have reshaped our understanding of hematopoiesis within the developing heart, highlighting its qualitative rather than quantitative importance.
Nevertheless, the dynamic nature of hematopoietic cells has made it challenging to definitively establish the presence, extent, and physiological relevance of endocardial hematopoiesis. Cre-based lineage tracing studies have struggled to capture this transient and spatially restricted process [15, 16]. However, the use of scRNA-seq has provided unprecedented resolution and revealed that endocardial cells possess an intrinsic hematopoietic program independent of other known hematopoietic organs [9, 13]. Emerging evidence from zebrafish models further supports the concept of endocardial hematopoiesis as a conserved phenomenon, reinforcing its developmental significance across species [17, 18].
Here we summarize the current knowledge on endocardial hematopoiesis, focusing on the conservation across species, the molecular mechanisms involved, its significance in vivo, as well as the challenges and future perspectives. Beyond endocardial hematopoiesis, this review summarizes studies showing the contribution of hematopoietic cells in cardiac development, providing new insights into the complexity of cardiogenesis.
Cardiogenic mesoderm-derived Hematopoiesis in DrosophilaIn Drosophila, signaling pathways involving bone morphogenetic protein (bmp), decapentaplegic (dpp), wingless (wg, Wnt1 homolog), and fibroblast growth factor (fgf) are essential for specifying the cardiac mesoderm, the common origin of cardioblasts, pericardial nephrocytes, and lymph gland hematopoietic progenitor cells [19, 20]. This developmental framework suggested a potential link between cardiogenesis and hematopoiesis, laying the groundwork for understanding how these processes are interconnected (Fig. 1a). While such relationships had been postulated, direct in vivo evidence demonstrating how a single progenitor cell divides into both blood and endothelial cells in vertebrates was lacking for a long time [5, 21,22,23].
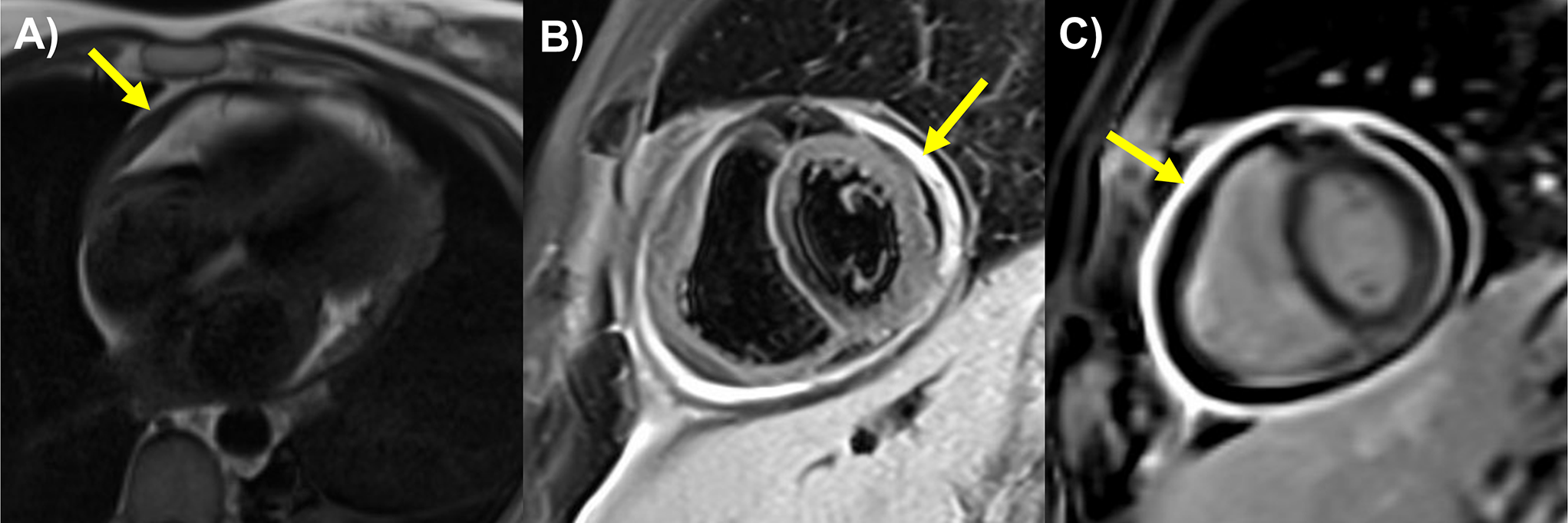
Fig. 1
Evolutionarily conserved mechanisms of common cardio-hematopoietic specification. (a) Drosophila cardiogenic mesoderm is the common origin of lymph gland hematopoietic progenitors, cardioblasts, and pericardial nephrocytes. The dynamic interaction between notch signaling and transcription factors such as srp, pnr, tin, and hand is essential for guiding the correct lineage commitment. (b) Embryonic hematopoiesis in zebrafish occurs in the ALPM, which serves as a common origin for both cardiac precursors and hemangioblasts during early embryonic stages. Recent studies have identified endocardial hematopoiesis around 24 hpf, primarily giving rise to neutrophils [16]. In later stages, endocardial hematopoiesis is also observed concurrently with definitive hematopoiesis, contributing to the formation of HSPCs [17]. (c) During mouse cardiogenesis, SHF-derived Nkx2-5-lineage endocardial cells undergo EHT through Notch signaling activation [6, 7]. Hemogenic endocardial cells (CD41+) differentiate into cardiac tissue macrophages (MΦs) by the inhibition of RA signaling through Dhrs3 expression, contributing to cardiac cushion remodeling and, consequently, valve maturation [7, 10]. Created in BioRender. Liu, N. (2025) https://BioRender.com/f04y383, and are partly adapted from Liu N and Nakano A, Front. Physiol. 2024 [14], under the terms of the Creative Commons Attribution License (CC BY)
Seminal studies in Drosophila larvae provided critical insights into the connection of cardiogenesis and hematopoiesis. Using the FLP/FRT system to trace labeled clones, researchers demonstrated that the cardiogenic mesoderm contributes to both lymph gland prohemocytes and cardioblasts [6]. This discovery was pivotal in identifying the dual potential of the cardiogenic mesoderm, a concept that would later resonate in vertebrate systems.
One of the key findings was the role of signaling pathways, particularly wg and notch, in regulating the differentiation of the cardiogenic mesoderm. Wg signaling promotes the differentiation of the mesoderm into both lymph gland and cardioblast lineages, whereas notch signaling acts as an inhibitor [6]. Notably, notch signaling functions in a time-dependent manner: early in development, it restricts the cardiogenic mesoderm, while later, it facilitates the production of lymph gland prohemocytes and reduces the formation of cardioblasts [6]. This temporal regulation highlights the complexity of mesodermal fate determination and suggests a fine-tuned molecular mechanism that ensures proper differentiation. At the molecular level, notch signaling regulates the expression of serpent (srp), a GATA transcription factor crucial for lymph gland differentiation [6]. In contrast, restricting notch signaling maintains the expression of tinman (tin, Nkx2-5 homolog) and pannier (pnr), key factors driving cardioblast formation [6]. This dynamic interaction between signaling pathways and transcription factors is essential for guiding the correct lineage commitment. The link between cardiogenesis and hematopoiesis is further reinforced by the discovery that the basic helix-loop-helix (bHLH) transcription factor hand acts as a common downstream target of both srp and tin [7]. This close lineage relationship between cardiognesis and hematopoiesis and the genetic regulatory mechanisms involved have been validated by more refined and recent lineage tracing experiments and scRNA-seq analyses [24, 25].
The conservation of these processes across species is further supported by the recent discovery of endocardial hematopoiesis in vertebrates. In species ranging from zebrafish to mammals, hematopoietic progenitors are derived from endocardium, a tissue derived from the cardiogenic mesoderm. This finding provides a direct link between cardiac progenitors and hematopoietic cells, mirroring the findings in Drosophila. The evolutionary conservation of this relationship suggests that the mechanisms governing the interaction between these two lineages are deeply rooted and likely played a critical role in the evolution of vertebrate circulatory systems.
Embryonic Hematopoiesis in ZebrafishLike the cardiogenic mesoderm in Drosophila, the ALPM in zebrafish harbors progenitors for both cardiac and hematopoietic lineages [26,27,28,29]. Within the ALPM, the rostral region contains hemangioblasts that give rise to hematopoietic, endothelial, and endocardial cells [30,31,32], while the adjacent posterior region comprises cardiac precursors that differentiate into cardiomyocytes [33, 34]. This spatial organization underscores the distinct gene regulatory programs that govern hemangioblast and cardiac progenitors, while also revealing their interplay (Fig. 1b). For example, mutant embryos of cloche, a gene essential for hematopoietic and endothelial specification [35], result in ectopic cardiomyocyte formation in the rostral region of the ALPM [36]. Likewise, knockout mice of hematopoietic transcription factor Scl/Tal1 activated cariac progenitor transcription in endothelial/endocardial cell lineages [37]. Moreover, transcription factors such as gata4, gata5, and gata6 act upstream of shared lineage-specific programs, highlighting the latent cardiogenic potential within the hemangioblast region [28, 30, 36, 38,39,40]. These findings suggest that the rostral ALMP region represents an evolutionary precursor to the mammalian secondary heart field (SHF), which is known to contribute to the outflow tract and arterial pole of the heart [28].
In zebrafish, SHF progenitors originating from the ALPM require conserved gene expression programs, including nkx2-5, which parallels mammalian SHF development [41]. Interestingly, the endocardium derived from the rostral ALPM not only contributes to cardiac morphogenesis but also exhibits hematopoietic potential. Time-lapse imaging and photoconversion experiments demonstrated endothelial-to-hematopoietic transition (EHT) in endocardial cells after 24 h post-fertilization (hpf) [17], coinciding with the incorporation of SHF progenitors into the heart [41]. This process is regulated by gata5/6 and the hedgehog signaling pathway, independent of etv2 and scl, which were previously thought to be master regulators of hematopoiesis, suggesting a new regulatory pathway specific to the endocardium [17]. Furthermore, this study performed scRNA-seq to reveale that endocardium-derived blood progenitor cells are mostly neutrophils, updating the previous recognition that hemangioblasts in the rostral region of the ALPM are a source of myeloid cells, including neutrophils [17]. These results suggest that endocardial hematopoiesis in zebrafish can be considered as a type of primitive hematopoiesis, since it is derived from ALPM and only contributes to myeloid cells.
In contrast, another study reported endocardial hematopoiesis in zebrafish embryos, which is reminiscent of definitive hematopoiesis. Bornhost et al. showed that endocardial cells derived from dorsal aorta endothelial cells undergoing EHT increase after 74 hpf, when cardiac looping is completed [18]. The study also used time-lapse imaging and Kaede photoconversion techniques, along with single-cell RNA-sequencing (scRNA-seq). Although the transcriptional regulatory mechanism of EHT was not determined, the scRNA-seq analysis revealed that most hemogenic endocardial cells express genes involved in promoting hematopoietic stem and progenitor cell (HSPC) fate in the caudal hematopoietic tissue (CHT), known as the definitive hematopoietic site in the zebrafish [18]. These endocardial cell-derived HSPCs are primarily localized in the outer curvature near the outflow tract, an SHF-derived region, and their adhesion is mediated by itga4 and vcam1 [18]. Moreover, HSPC production from endocardial cells was suggested to be involved in the maintenance of systemic erythrocyte counts in a compensatory manner [18]. Taken together, the study showed that the endocardium contributes to hematopoiesis both as a tissue source and as a niche, possibly reproducing definitive hematopoiesis.
Although the molecular mechanisms and functional significance of these processes remain to be fully elucidated, the zebrafish endocardium has been shown to harbor hematopoietic programs resembling both primitive and definitive hematopoiesis. Notably, each program shares certain similarities with mammalian endocardial hematopoiesis. Given that the mechanisms underlying endocardial hematopoiesis in mammals remain a topic of ongoing debate, these findings in zebrafish offer a valuable framework for exploring the conserved and divergent aspects of endocardial hematopoiesis across species. As such, insights from zebrafish studies may serve as a foundation for developing new models to better understand the role of the endocardium in mammalian hematopoiesis, shedding light on its evolutionary and developmental significance.
Endocardial Hematopoiesis in MouseIn mammals, hemangioblasts are present as yolk sac blood islands, which are functionally analogous to those described in Drosophila and zebrafish. These blood islands give rise to both endothelial and hematopoietic progenitor cells, playing essential roles in primitive hematopoiesis and vasculogenesis. Within the embryo proper, however, distinct hemangioblasts with hemo-endothelial biopotency have not been identified. Instead, endothelial cells in regions such as the AGM undergo EHT, acquiring hematopoietic potential and generating hematopoietic progenitor cells.
Interestingly, while cardiogenic progenitors in Drosophila and zebrafish emerge from primitive hematopoietic regions such as the cardiogenic mesoderm and ALPM, mammalian cardiogenic progenitors in the lateral plate mesoderm of the embryo proper exhibit a unique feature. During the very early stages of cardiac progenitor specification, these mammalian cardiac progenitors transiently express hematopoietic transcription factors [42], reflecting a potential evolutionary connection between hematopoiesis and cardiogenesis. Although latent hematopoietic potential in cardiac progenitor cells was suggested transcriptionally more than 2 decades ago, bona fide hematopoiesis from the cardiac region has not been examined for a long time. Because of its scarcity and transient nature, hematopoiesis from the heart has long been overlooked, but our close observation and the use of scRNA-seq analysis (GSE126128 [43], GSE76118 [44]), we have captured that expression of hematopoietic transcription factors is maintained in a subset of endocardial cells during cardiac formation [6, 7] The details of the discovery and proof of endocardial hematopoiesis in mice are described below.
The plasticity of endocardial cells has predominantly been examined in relation to their roles in mesenchymal cell differentiation in the cardiac cushion and the formation of coronary endothelial cells. However, their potential to contribute to hematopoiesis remains an underexplored area [45,46,47,48,49]. Endocardial cells, which line the heart’s inner surface and typically exhibit a squamous morphology, undergo a marked transformation during hematopoietic induction. These cells adopt a rounded shape and begin expressing early hematopoietic markers such as CD41 and Tal1 [9, 11]. Both our findings and those of others have revealed the presence of endocardial cells expressing hematopoietic markers within specific regions of the mouse embryonic heart, including the outflow tract, atrioventricular canal, and inflow tract [9, 12]. This endocardial hematopoiesis is spatiotemporally coincident with endocardial cushion formation from SHF-derived endocardium, which is regulated by Nkx2-5 (Fig. 1c) [6, 49, 50]. Nkx2-5 knockout mice, which fail to develop endocardial cushions and exhibit hypoplastic cardiomyocytes, die during mid-gestation [50, 51]. Importantly, these mice also exhibit severe hematopoietic defects in both the yolk sac and the endocardium, highlighting the multiple roles of Nkx2-5 in regulating EHT, endothelial-to-mesenchymal transition (EndoMT), and cardiogenesis in mammals [9, 50, 51]. Of note, endocardial hematopoiesis occurs independently of regulation by Runx1, suggesting a primitive nature of the endocardial hematopoiesis [9, 13].
The scRNA-seq analysis (GSE76118 [44]) identified two critical pathways involved in Nkx2-5-dependent endocardial hematopoiesis: Notch signaling and retinoic acid (RA) signaling [10]. Activation of Notch signaling in Nkx2-5-lineage cells enhanced endocardial hematopoiesis [10]. This finding aligns with tin/notch hematopoiesis in Drosophila cardiogenic mesoderm [6], suggesting an evolutionarily conserved role for endocardial hematopoiesis. Importantly, activation of Notch signaling in Nkx2-5 lineage cells in Nkx2-5 null-background mice restored the Nkx2-5 knockout endocardial phenotype, such as lack of endocardial cushion and endocardial hematopoiesis [10]. This suggests that Notch signaling plays a pivotal role in regulating the bilateral differentiation of endocardial cells into mesenchymal and hematopoietic cells by Nkx2-5/Notch signaling, highlighting the mutual regulation of cardiac and hematopoietic development. In zebrafish, notch signaling also governs endocardial fate decisions, driving endocardial cushion formation by directly upregulating EndoMT genes [52, 53]. Additionally, it regulates EHT in the ventral dorsal aorta, promoting HSC emergence [54]. These findings underscore the conserved role of Notch signaling in shaping endocardial cell fate across species.
The involvement of RA in endocardial hematopoiesis was identified through a ligand-receptor-target gene network analysis, called NicheNet [55] using scRNA-seq data [10]. The enzyme gene Dhrs3 (Dehydrogenase/reductase 3), which regulates the reduction of all-trans retinoic acid (atRA), is one of the most targeted genes in the hemogenic endocardial cell cluster and is significantly downregulated in Nkx2-5 knockout endocardial cells [10]. Ex vivo hematopoietic colony formation assays revealed that excessive RA signaling impaired the differentiation of hematopoietic progenitors, including macrophages, underscoring the importance of RA suppression via Dhrs3 expression in endocardial hematopoiesis [10].
Taken together, during mouse cardiogenesis, SHF-derived Nkx2-5-lineage end
Comments (0)