Hormones play a crucial role in shaping the development and behaviour of organisms across the animal kingdom. In insects, juvenile hormone (JH), through its intracellular receptor, Met/Gce (Wilson and Fabian, 1986, Charles et al., 2011, Jindra et al., 2015, Tumova and Jindra, 2023), is a central regulatory molecule, influencing various aspects of an individual's life (Riddiford, 2020), from its development and reproduction (Riddiford, 2012, Barton et al., 2024) to caste differentiation (Corona et al., 2016) and division of labour, and foraging behaviour in social insects (Elekonich et al., 2001, Norman and Hughes, 2016).
JH is synthesized from acetyl-CoA by the cells of cephalic endocrine glands, the corpora allata, in honeybees (Rachinsky and Hartfelder, 1990), through a series of enzymatic reactions (universal eukaryotic mevalonate pathway), which have been extensively described (e.g. Bellés et al., 2005, Tsang et al., 2020). The first molecular precursor within the mevalonate pathway branch leading to JH synthesis (the other branch leads to cholesterol synthesis and is absent in insects) is farnesol. Farnesol is first oxidated to farnesal (by farnesol oxidase), then dehydrogenated to farnesoic acid (by farnesal dehydrogenase), methylated to methyl farnesoate (by JH methyltransferase), and, finally, epoxidated (by farnesoate epoxidase) to JH (Bellés et al., 2005).
The last steps to JH production (from farnesoic acid) have evolved differently in different insect orders (Tsang et al., 2020) and may also take place in peripheral tissues (e.g. Borovsky et al., 1994, Shinoda and Itoyama, 2003). Additionally, the last precursors have been reported to have per se regulatory function in development, metamorphosis, and reproduction (Cheong et al., 2015, De Loof et al., 2015). Pioneer works on the activity of purified compound in Rodnius (Schmialek, 1961., Wigglesworth, 1961) and on functional screening using Galleria wax test already showed the diverse characteristics of JH-like activity of farnesyl derivatives and related compounds (Schneiderman et al., 1965). Importantly, as recently shown, epoxidation of JH precursors (the last step) seems to have played a key evolutionary role providing a reproductive advantage to insects (Nouzova et al., 2021).
An additional level of complexity in the function of JH-related compounds in insect biology raises within the context of insect-plant interaction. Plants usually produce secondary substances they use to attract or repel numerous insects (Nishida, 2014). Some of these substances are JH-related compounds and unraveling how they modulate insect’s physiology and behaviour is paramount to understand the ecology and evolution of insect groups. Advances in this field were achieved in recent years. For instance, reports suggested that nerolidol (a natural sesquiterpenol with structural similarities to JHs) can function as a JH analogue by modulating the expression of the enzyme genes responsible for degrading JH, resulting in JH disorders, and ultimately disrupting the development of insect larvae (Dai et al., 2023). Moreover, several JH agonists and antagonists causing retardation in the ovarian development of female Asian tiger mosquito, Aedes albopictus, were identified from plant essential oil compounds (Lee et al., 2018), and others have shown to alter the insect’s gene expression patterns and seem to be species-specific (Lee et al., 2015, Shin et al., 2018).
Farnesol, a naturally occurring sesquiterpene alcohol found in all eukaryotes, has attracted attention due to its diverse biological activities (De Loof et al., 2015). In addition to being a JH precursor in insects, it is involved in signalling, communication, and antimicrobial defence (Polke et al., 2018) and has been early identified as a receptor ligand, activating the vertebrate FXR-RXR heterodimer receptor (Forman et al., 1995). Farnesol's unique role as a quorum-sensing molecule in bacteria and yeast is well-documented, regulating gene expression and behaviours in response to population density (Leonhardt et al., 2015, Dižová and Bujdáková, 2017). Farnesol is a constituent of floral volatiles (e.g. Wu et al., 2021, Li et al., 2022), suggesting its importance in pollination and foraging behaviour. Furthermore, farnesol is found in the resin of Baccharis dracunculifolia L. (Asteraceae, Astereae, Baccharidinae) (Alencar et al., 2005), popularly known in Brazil as “alecrim-do-campo” or “vassourinha” and in Argentina as “chilca”. B. dracunculifolia plants were documented in South America, mainly in Southwest Brazil, Argentina, Paraguay, and Uruguay (Heiden, 2021, Olivares-Pinto et al., 2022). Honeybees (Apis mellifera) are fond of B. dracunculifolia, from which they get resin and elaborate the most worldwide marketable propolis, green propolis (Pereira et al., 2009, Bastos et al., 2011). As expected, significant amounts of farnesol were also detected in this propolis (Teixeira et al., 2005), which bees use to seal cracks in the walls, reinforce the wax combs, and act against bacteria and fungi (Seeley, 1995, Wilson et al., 2013). This wide range of roles played by farnesol in different contexts raises intriguing questions about its potential involvement in honeybee physiology, development, and behaviour, particularly in the context of hormone-like effects. This because farnesol can be brought to the nest by foraging honeybee workers and spread into the colony members (including immatures) and through the nesting material.
The introduction within a honeybee hive of a compound with potential hormonal activity could be eventually a challenge to the colony survival, mainly because the division of labour among workers is under hormonal control. Briefly, the division of labour is based on a pattern of behavioural development in which workers perform different tasks as they age. Bees perform within-hive activities such as caring for brood (nursing) and cleaning for the first 2 to 3 weeks of adult life and then they start performing outside activities mainly foraging nectar and pollen for 1 to 2 weeks until they die (Seeley, 1982). The transition is promoted and paced by JH (Robinson, 1987, Sullivan et al., 2000, Robinson and Vargo, 1997) and is critical for the colony's survival and productivity, as it ensures that the necessary tasks are carried out by bees with the appropriate age-related traits.
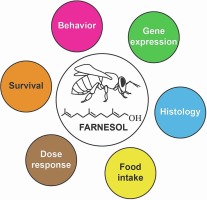
Here, we tested the hypothesis that exogenous farnesol alters the pacing of developing workers. This effect could be reached by farnesol acting as a hormone per se or as a modified precursor of JH (Chai and Shuel, 1970, Bendena et al., 2011), thus increasing endogenous JH levels. In a series of laboratory assays, we first tested farnesol effects in the diet of young honeybee workers and found profound alterations in molecular, morphological, and behavioural parameters. Nevertheless, since the results could be due to the observed higher levels of food ingestion in the treated bees, we next performed similar tests by topically applying farnesol during pharate-adult development of workers, and then, in young honeybees. Our results showed exogenous farnesol has JH-like effects in developing and young honeybee workers, likely after being used as a JH precursor.


留言 (0)